|
| |

Marsupiaux éteints récemment
|
Nom |
Statut |
Parcours et cause
disparition (EX) |
Thylacine
(Thylacinus cynocephalus)
(Moins de 130 cm, moins de 30 kg)

 |
EX
|
Origine et évolution Le thylacine (Thylacinus
cynocephalus), aussi appelé tigre de Tasmanie ou loup marsupial, était un
mammifère carnivore marsupial appartenant à l’ordre des Dasyuromorphia. Il
ne partageait aucun lien évolutif direct avec les canidés malgré sa
ressemblance morphologique, fruit d’une évolution convergente. Son lignage
remonte au Miocène, avec plusieurs espèces fossiles apparentées. Le
thylacine était le dernier représentant de la famille des Thylacinidae,
aujourd’hui éteinte. Des études génétiques ont révélé une faible diversité
génétique bien avant l’arrivée des humains en Australie, ce qui aurait
compromis sa résilience évolutive.
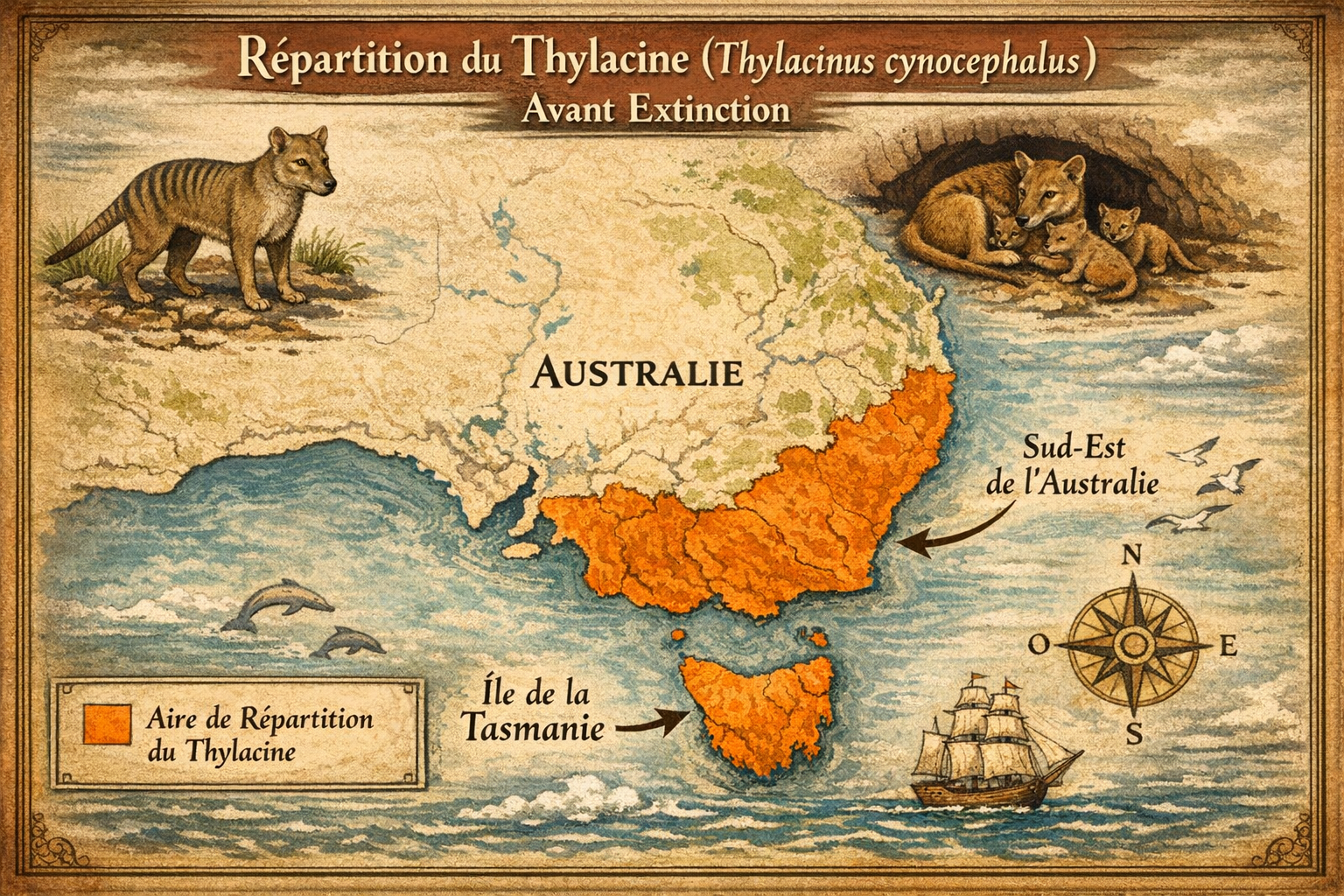
Répartition et écologie Autrefois largement répandu en Australie
continentale et en Nouvelle-Guinée, le thylacine a vu son aire se réduire à
la seule île de Tasmanie il y a environ 3000 ans, probablement à cause de la
compétition avec le dingo. Il occupait des habitats variés, notamment les
forêts et les zones ouvertes. Prédateur solitaire, nocturne ou
crépusculaire, il se nourrissait de kangourous, wallabies, oiseaux nichant
au sol et petits mammifères. Il possédait une poche marsupiale et un pelage
rayé distinctif sur le dos, d’où son surnom de tigre de Tasmanie.
Causes de la disparition La disparition du thylacine est
principalement due à la chasse intensive encouragée par des primes
gouvernementales dès le XIXe siècle, les colons le considérant comme une
menace pour le bétail. L’introduction de chiens domestiques, la perte
d’habitat, les maladies et la faible diversité génétique ont aggravé son
déclin. Le dernier individu connu est mort en captivité au zoo de Hobart en
1936. Des observations non confirmées ont été rapportées depuis, mais aucune
preuve scientifique n’a validé leur véracité
Confirmation de l’extinction L’UICN classe officiellement le
thylacine comme espèce éteinte depuis 1982. Malgré des recherches et des
signalements sporadiques en Tasmanie et en Papouasie-Nouvelle-Guinée, aucune
preuve tangible n’a été apportée. Des projets de désextinction sont en
cours, notamment à l’université de Melbourne, visant à recréer une forme
génétiquement proche à partir d’ADN ancien, mais ces initiatives restent
controversées et expérimentales |
Chaeropus
ecaudatus (Bandicoot à pieds de cochon)
(Moins de 30 cm, moins de 300 g)

 |
EX
|
Origine et évolution Chaeropus ecaudatus, appelé bandicoot
à pieds de cochon, était un petit marsupial australien appartenant à la
famille aujourd’hui éteinte des Chaeropodidae. Il représentait un genre
unique, Chaeropus, distinct des autres bandicoots par sa morphologie
singulière. Son évolution a produit des adaptations inhabituelles, notamment
des pieds antérieurs avec deux orteils fonctionnels dotés de sabots, et des
pieds postérieurs avec un seul orteil locomoteur en forme de petit sabot de
cheval. Ces traits évoquent une convergence évolutive avec des ongulés, bien
qu’il s’agisse d’un marsupial.
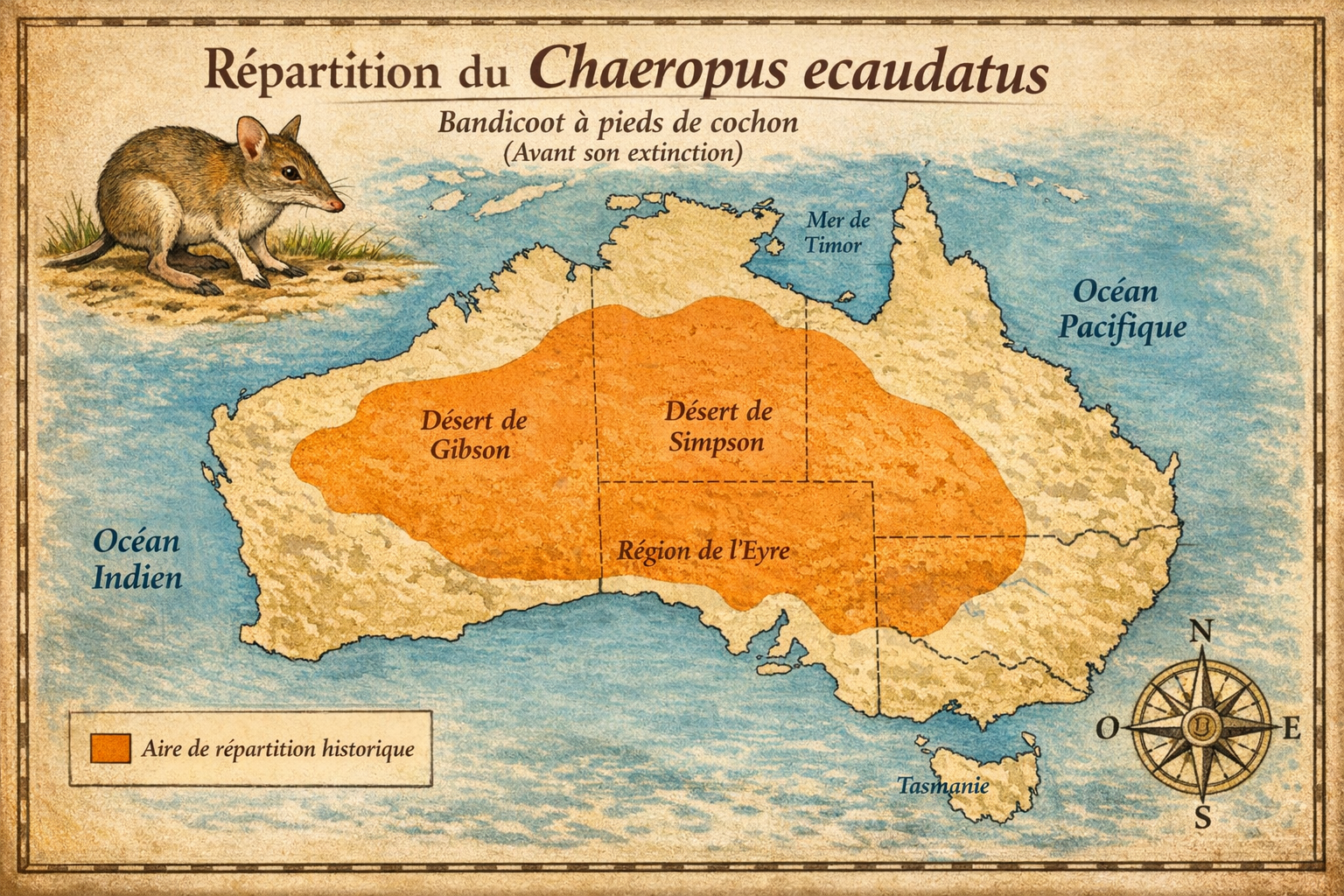
Répartition et écologie L’espèce était présente dans les plaines
arides et semi-arides de l’intérieur de l’Australie, notamment dans le
nord-ouest du Victoria, l’Australie-Méridionale, l’Australie-Occidentale et
le Territoire du Nord. Elle occupait divers habitats allant des prairies
herbeuses aux zones à spinifex. Les témoignages aborigènes indiquent qu’il
construisait des nids d’herbe pour dormir le jour et pouvait fuir à grande
vitesse s’il était dérangé. Il creusait aussi des terriers courts et
rectilignes. Herbivore, il se nourrissait probablement de végétaux secs
adaptés aux milieux désertiques.
Causes de la disparition Le bandicoot à pieds de cochon a souffert
de la modification de son habitat par le pâturage intensif, de la prédation
par les espèces introduites comme les renards et les chats, et de maladies
potentiellement transmises par le bétail. Sa faible densité et sa discrétion
ont rendu difficile sa détection et sa protection. Le dernier spécimen connu
a été collecté en 1901, bien que l’espèce ait été officiellement considérée
comme éteinte seulement en 2004.
Confirmation de l’extinction L’UICN classe Chaeropus ecaudatus
comme espèce éteinte. Aucun individu vivant n’a été observé depuis le début
du XXe siècle. Les rares spécimens conservés dans les musées et les
témoignages aborigènes constituent les seules sources d’information sur son
comportement. Sa disparition illustre la fragilité des espèces endémiques
face aux bouleversements écologiques rapides et non maîtrisés |
Caloprymnus campestris (Kangourou-rat du désert)
(Moins de 25 cm, moins de 200 g)

|
EX
|
Le Caloprymnus campestris, ou kangourou-rat du désert, a disparu
officiellement en 1935, victime de la dégradation de son habitat et de
l’introduction de prédateurs exogènes.
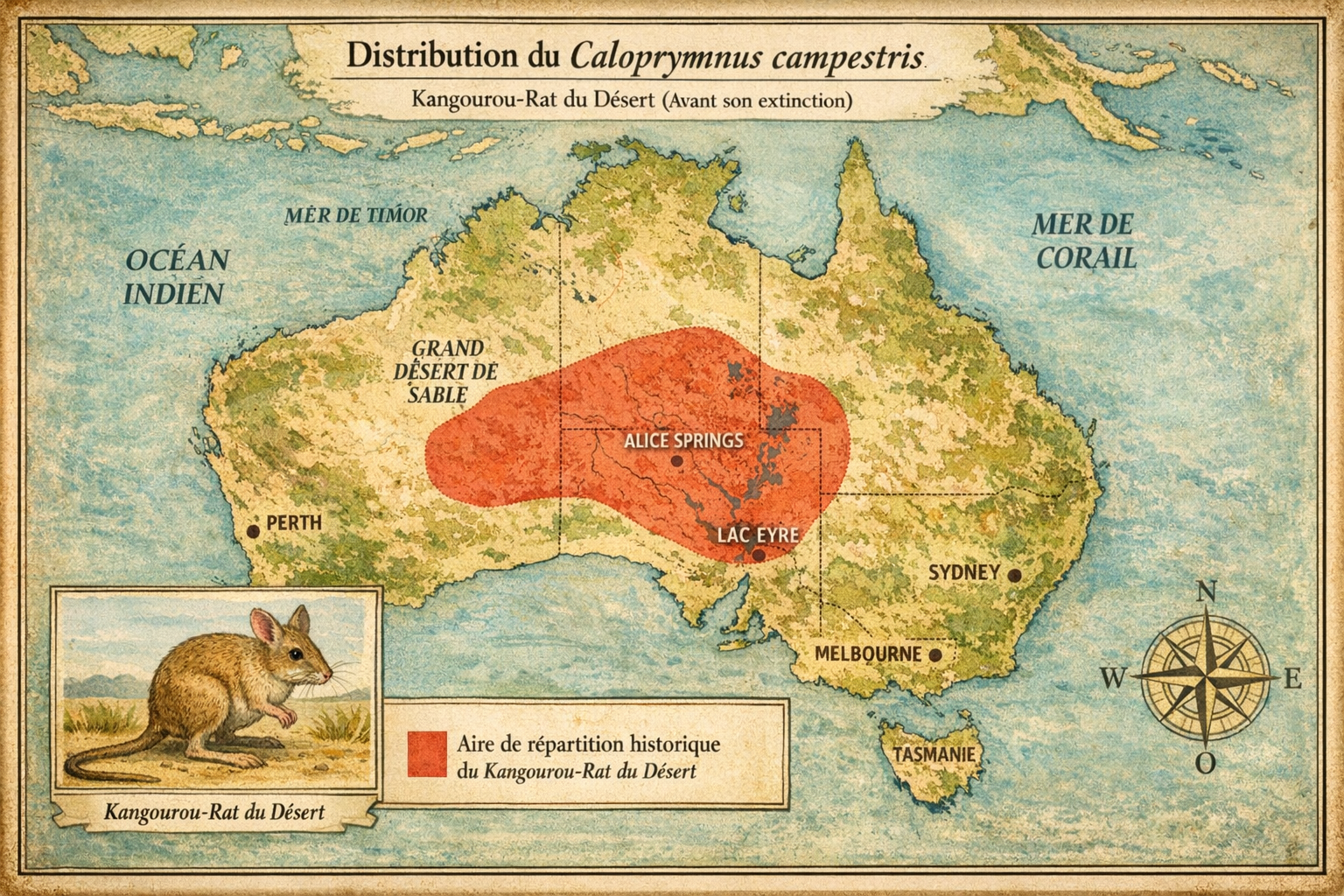
Découverte et répartition initiale L’espèce est découverte en 1841
dans le centre de l’Australie, notamment dans les zones arides du Sturt
Stony Desert, du Birdsville Track et du Bunda Plateau. Elle est décrite par
John Gould en 1843 à partir de spécimens envoyés par George Grey. Le
Caloprymnus campestris vivait dans les regs, les dunes et les sols argileux,
construisant des abris rudimentaires sous les buissons ou à l’air libre. Il
était endémique d’Australie centrale et jamais abondant.
Observations et raréfaction Après sa découverte, l’espèce n’est
plus observée pendant plusieurs décennies. Des témoignages non confirmés
apparaissent entre 1876 et 1905, notamment près de Birdsville et du lac
Killalpaninna. En 1935, Hedley Finlayson photographie et collecte les
derniers spécimens connus. Des observations non vérifiées sont rapportées
après les pluies de 1956-1957 et 1974-1975, mais aucune preuve tangible ne
permet de confirmer leur validité.
Causes de la disparition La disparition du Caloprymnus campestris
est attribuée à plusieurs facteurs convergents. La dégradation de son
habitat par le surpâturage, les feux mal gérés et l’exploitation humaine a
réduit ses zones de refuge. L’introduction de prédateurs comme le renard
roux et le chat domestique a accentué la pression sur une population déjà
fragile. La compétition avec les lapins pour les ressources alimentaires a
aggravé la situation. L’espèce, peu mobile et spécialisée, n’a pas pu
s’adapter à ces bouleversements rapides.
Statut actuel et perspectives L’espèce est classée comme éteinte
par l’UICN depuis 1994. Toutefois, des recherches récentes sur sa
morphologie crânienne et ses habitudes alimentaires suggèrent qu’elle
pourrait survivre dans des zones reculées du désert australien. Des
campagnes de suivi basées sur ces données sont envisagées pour tenter une
redécouverte. |
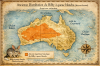
Macrotis
leucura (Bilby à queue blanche)
(Moins de 45 cm, moins de 1,5 kg)

 |
EX
|
Le Macrotis leucura, ou bilby à queue blanche, a disparu
officiellement dans les années 1960, victime de la prédation par des espèces
introduites et de la modification de son habitat désertique.
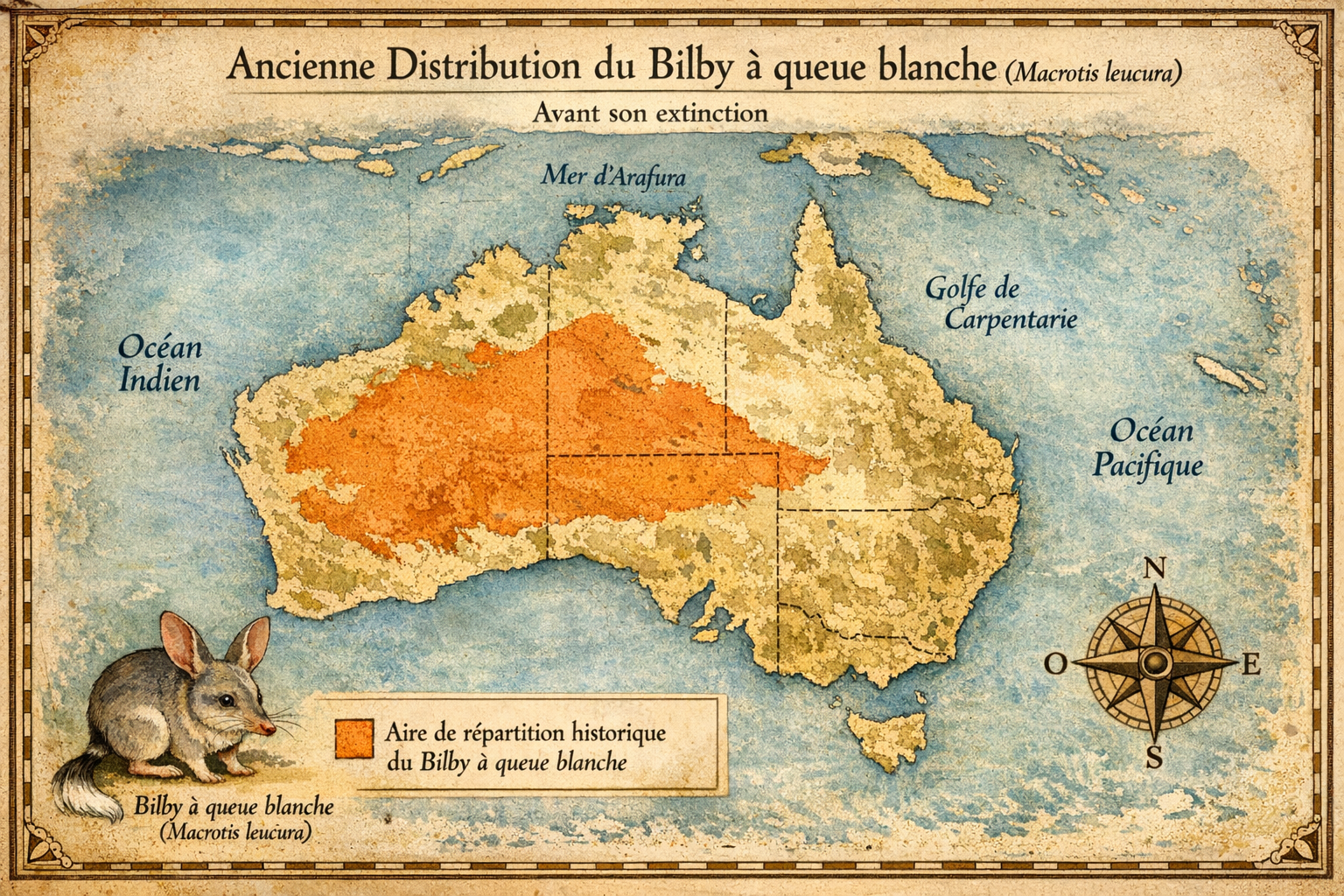
Découverte et répartition initiale L’espèce est décrite en 1887 par
Oldfield Thomas. Elle occupait les déserts de sable du centre de l’Australie
: désert de Gibson, Grand Désert de Sable, Simpson Desert, et zones arides
du Territoire du Nord, d’Australie-Occidentale et d’Australie-Méridionale.
Elle creusait des terriers profonds en spirale, obstrués à l’entrée, dans
les dunes et les plaines sableuses.
Morphologie et mode de vie Plus petit que le bilby actuel, le
Macrotis leucura mesurait entre 32 et 44 cm, avec une queue blanche de 11 à
27 cm. Il possédait trois doigts aux pattes arrière, dont deux fusionnés.
Nocturne et solitaire, il se nourrissait d’insectes, de petits mammifères,
de graines et de fruits, notamment du genre Solanum. Il ne buvait pas d’eau,
tirant son hydratation de son alimentation. La femelle portait huit mamelles
et donnait naissance à un à trois petits après une gestation de 21 jours.
Observations et raréfaction Les derniers spécimens vivants sont
observés en 1931 à Koonchera Dune. Un crâne est retrouvé en 1967 dans un nid
d’aigle au sud-est d’Alice Springs, suggérant une survie jusqu’au début des
années 1950. Des témoignages aborigènes évoquent sa présence jusqu’en 1960
dans certaines zones reculées.
Causes de la disparition La principale cause est la prédation par
le renard roux et le chat domestique, introduits par les colons européens.
Ces prédateurs ont décimé les populations de petits marsupiaux dans les
déserts australiens. La disparition des pratiques traditionnelles de brûlage
par les communautés aborigènes a modifié les régimes de feu, réduisant la
diversité végétale et les refuges. L’espèce, déjà rare et peu étudiée, n’a
pas résisté à ces pressions combinées.
Statut actuel et perspectives Classé comme éteint par l’UICN, le
Macrotis leucura n’a pas été revu depuis les années 1960. Aucun programme de
réintroduction n’est envisageable faute de spécimens vivants. Il reste un
symbole de l’impact des espèces invasives sur la faune australienne. |
Siréniens éteints récemment
|
Nom |
Staut |
Parcours et cause
disparition (EX) |
Rhytine
de Steller (hydromalys gigas)
(moins de 9 m, mois de 6t)
 
|
EX |
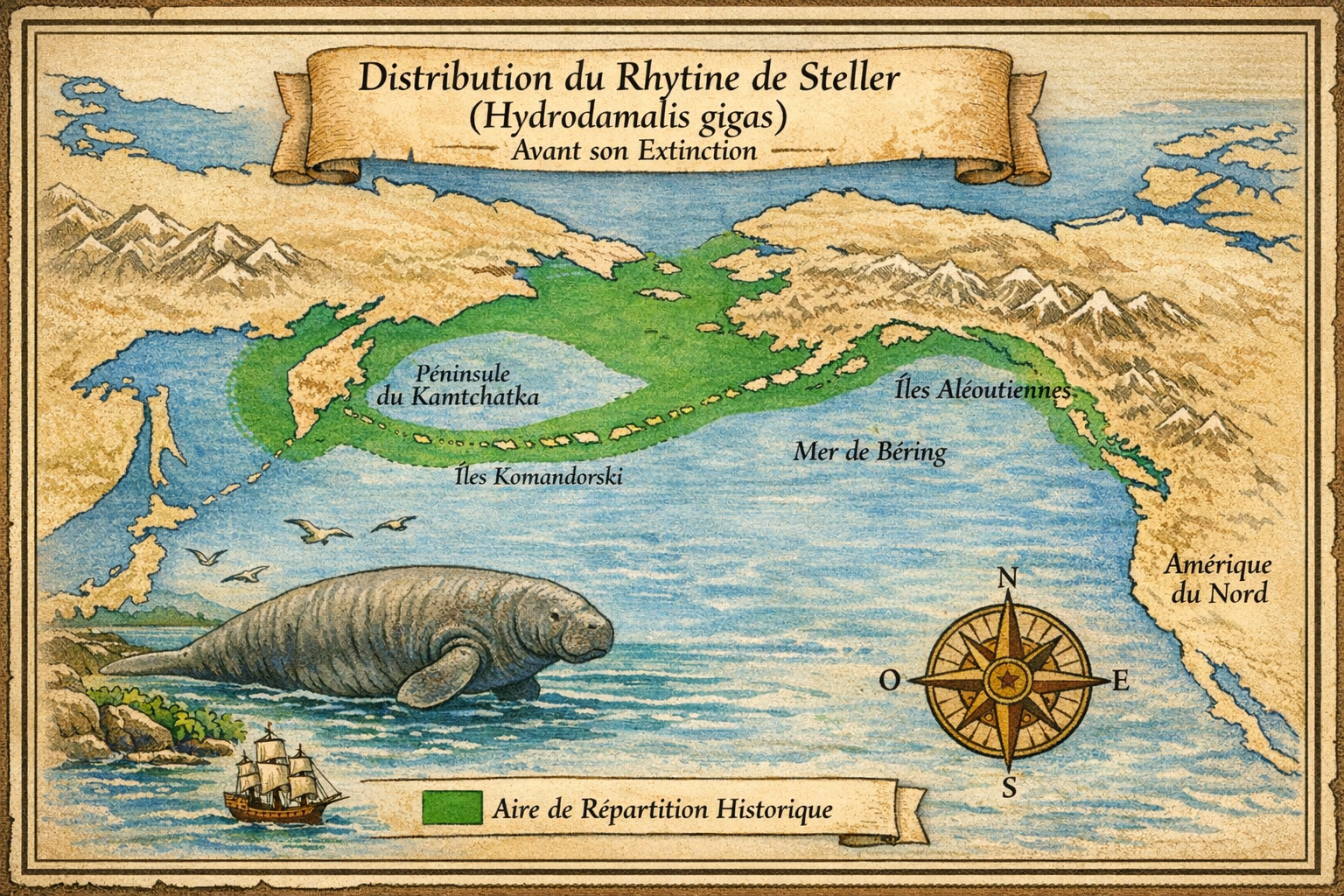
La Rhytine de Steller (Hydrodamalis gigas) a disparu en 1768,
seulement 27 ans après sa découverte, victime d’une chasse intensive et de
son extrême vulnérabilité comportementale.
Découverte et répartition initiale L’espèce est découverte en 1741
par Georg Wilhelm Steller lors de l’expédition de Vitus Béring dans la mer
de Béring. Elle vivait exclusivement autour des îles du Commandeur (Béring
et Medny), bien que des fossiles indiquent une répartition historique plus
large sur le pourtour du Pacifique nord, du Japon à la Californie.
Morphologie et écologie La Rhytine mesurait entre 8 et 9 mètres
pour un poids de 8 à 10 tonnes. Elle appartenait à la famille des dugongidés,
mais se distinguait par sa taille massive, sa peau noire épaisse et plissée,
et sa flottabilité positive qui l’empêchait de s’immerger complètement. Elle
se nourrissait exclusivement d’algues marines et vivait en groupes
familiaux. Sa gestation était longue et sa reproduction lente, ce qui la
rendait particulièrement vulnérable.
Causes de la disparition La disparition est due à une chasse
systématique dès le retour de l’expédition Béring. Les marins et marchands
de fourrure exploitent sa graisse, sa viande, son lait et sa peau. Son
comportement placide, son absence de peur de l’homme et sa lenteur ont
facilité son extermination. En l’absence de prédateurs naturels et de
mécanismes de défense, la population estimée à 2 000 individus est éradiquée
en moins de trois décennies.
Statut actuel et postérité L’espèce est classée comme éteinte par
l’UICN. Elle représente l’un des cas les plus rapides d’extinction
post-découverte chez les mammifères. Des témoignages non vérifiés ont
circulé au XIXe siècle, mais aucune preuve scientifique n’a confirmé leur
validité. La Rhytine de Steller est aujourd’hui un symbole de l’impact
humain sur les écosystèmes marins arctiques. |
Rongeurs éteints récemment
|
Nom |
Statut |
Parcours et cause
disparition (EX) |
Melomys
rubicola — Rat de Bramble Cay
(moins de 20 cm, mions de 120 g)

 |
EX
|
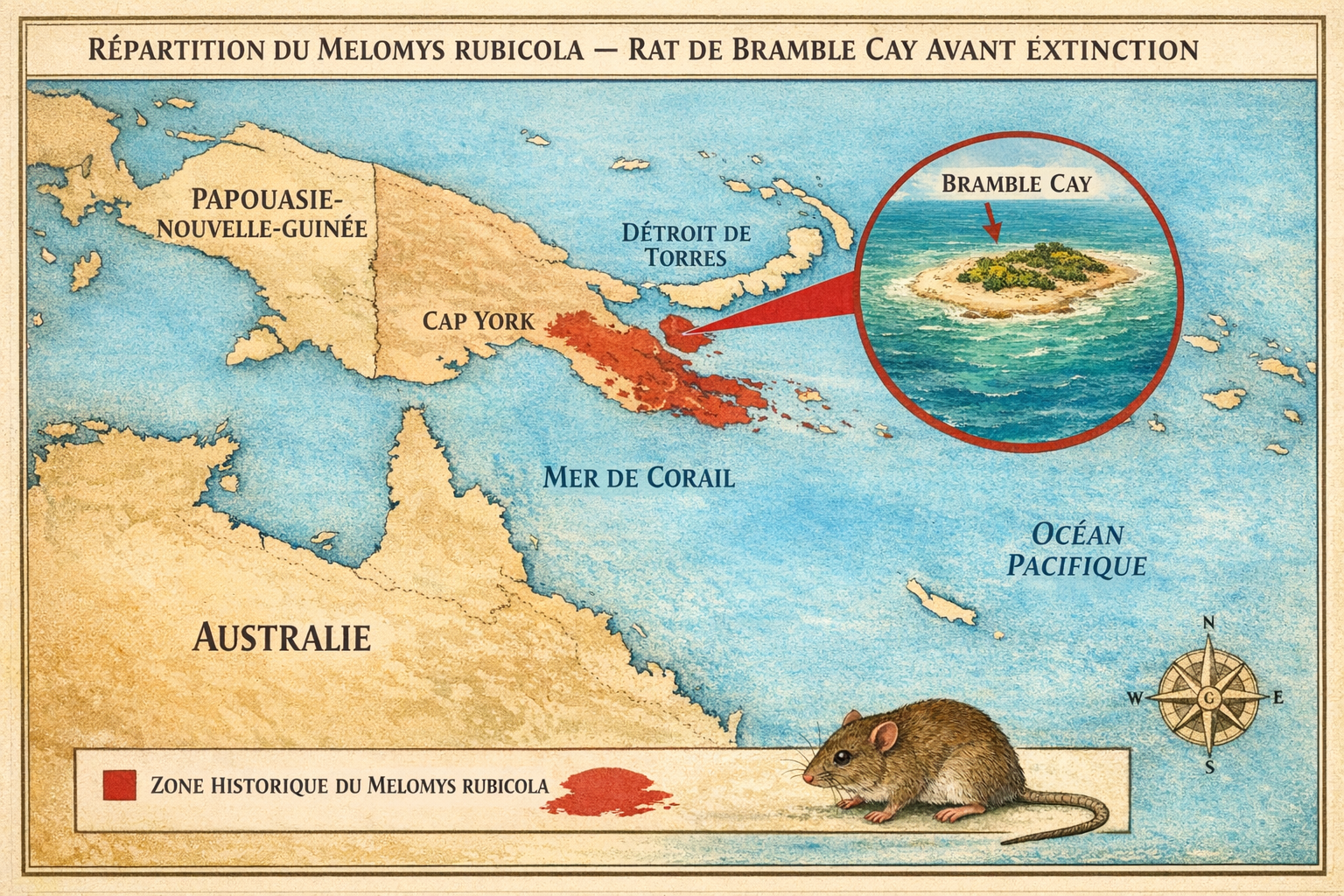
Le Melomys rubicola, ou rat de Bramble Cay, est le premier mammifère
officiellement déclaré éteint à cause du changement climatique anthropique.
Découverte et répartition initiale L’espèce est découverte en 1845
sur Bramble Cay, un îlot corallien situé à l’extrémité nord de la Grande
Barrière de corail, dans le détroit de Torrès. Elle est décrite
scientifiquement en 1924 par Oldfield Thomas. Endémique de cette île de 340
mètres sur 150, elle n’a jamais été observée ailleurs, malgré des recherches
sur le continent papou à 50 km de distance.
Morphologie et écologie Ce rongeur possédait une fourrure brun
rougeâtre sur le dos et grisâtre sur le ventre, une queue préhensile à
écailles en mosaïque, et mesurait environ 15 cm pour un poids de 80 à 160 g.
Il se nourrissait de végétation herbacée et vivait dans les zones les plus
élevées de l’île, à peine 3 mètres au-dessus du niveau de la mer.
Observations et raréfaction L’espèce était encore modérément
commune dans les années 1920. Elle est photographiée en 2001, mais aucune
observation n’est enregistrée après 2009. Une enquête exhaustive menée en
2014 par le gouvernement du Queensland et l’Université du Queensland
confirme l’absence totale de spécimens vivants. L’extinction est
officiellement reconnue en 2016 par l’UICN et en 2019 par le gouvernement
australien.
Causes de la disparition La cause principale est la montée du
niveau de la mer, qui a submergé l’habitat et détruit plus de 90 % de la
végétation de Bramble Cay entre 2004 et 2014. Les tempêtes tropicales et les
inondations répétées ont aggravé la situation. L’espèce, confinée à une zone
minuscule sans possibilité de migration, n’a pas pu survivre à cette perte
d’habitat. Aucun prédateur ou facteur humain direct n’est impliqué, ce qui
rend cette extinction emblématique du changement climatique.
Statut actuel et postérité Le Melomys rubicola est considéré comme
le premier mammifère dont l’extinction est directement liée à l’impact
climatique humain. Il est devenu un symbole de la vulnérabilité des espèces
insulaires face à la montée des océans et à la perte d’habitat causée par
les dérèglements climatiques. |
Megalomys
desmarestii — Rat géant de la Martinique
(moions de 40 cm, moins de 1,5 kg)

 |
EX
|
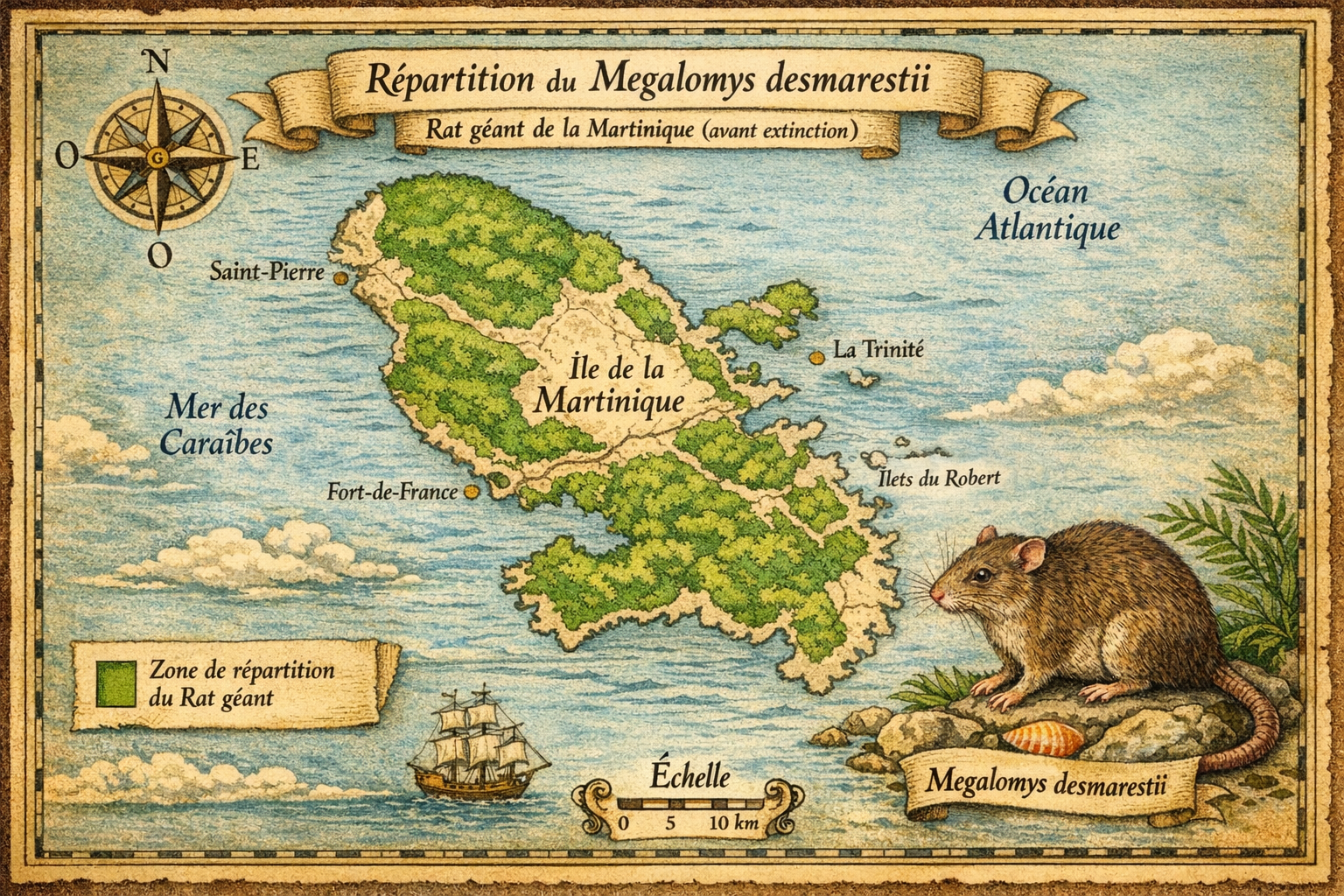
Le Megalomys desmarestii, ou rat géant de la Martinique, a disparu
au début du XXe siècle, victime de la prédation par des espèces introduites
et de bouleversements écologiques majeurs.
Découverte et répartition initiale L’espèce est décrite en 1829 par
Johann Fischer. Elle était endémique de la Martinique et représentait l’un
des plus grands rongeurs des Petites Antilles. Elle occupait divers habitats
de l’île, des zones côtières aux forêts d’altitude, avec une préférence pour
les milieux humides et boisés.
Morphologie et écologie Ce rongeur pouvait atteindre 30 à 40 cm de
long, avec une queue presque aussi longue que le corps. Il possédait une
fourrure dense, une tête massive et des incisives puissantes. Il se
nourrissait de végétaux, de fruits et de racines. Les populations
amérindiennes et esclaves le consommaient, malgré une odeur forte qui
nécessitait une cuisson prolongée et un assaisonnement spécifique.
Raréfaction et dernières observations Au XIXe siècle, l’espèce
devient rare, se repliant dans les forêts d’altitude sur les contreforts de
la montagne Pelée. Aucune observation confirmée n’est enregistrée après
l’éruption de 1902, qui détruit Saint-Pierre et bouleverse les écosystèmes
du nord de l’île.
Causes de la disparition La principale cause est l’introduction du
rat noir (Rattus rattus) par les colons européens, qui entre en compétition
pour les ressources et transmet des maladies. L’arrivée de la mangouste
indienne (Herpestes javanicus) pour lutter contre les serpents et les rats
accentue la pression prédatrice. L’éruption de la montagne Pelée en 1902,
bien que secondaire, a pu achever les dernières populations isolées.
Statut actuel et postérité L’espèce est classée comme éteinte par
l’UICN. Elle illustre les effets combinés de la bioturbation coloniale, de
la prédation invasive et des catastrophes naturelles sur les espèces
insulaires. Elle fait partie d’un groupe de rongeurs géants antillais
aujourd’hui disparus, dont Megalomys audreyae (Barbade) et Megalomys
curazensis (Curaçao). |
Megalomys
luciae — Rat géant de Sainte-Lucie
(moins de 30 cm, moins 1,2 kg)

 |
EX
|
Le Megalomys luciae, ou rat géant de Sainte-Lucie, a disparu au
milieu du XIXe siècle, victime de la prédation invasive et de la
colonisation écologique.
Découverte et répartition initiale L’espèce est décrite en 1901 par
Forsyth Major à partir de spécimens collectés sur l’île de Sainte-Lucie.
Elle était endémique de cette île des Caraïbes et représentait l’un des plus
grands rongeurs insulaires antillais. Elle occupait les zones boisées et
humides, probablement en altitude, mais son écologie reste mal documentée.
Morphologie et écologie Ce rongeur possédait une taille imposante,
comparable à celle d’un gros rat musqué, avec une queue longue et
écailleuse. Il se nourrissait de végétaux, de fruits et de racines. Son
comportement était probablement nocturne et discret, ce qui a limité les
observations directes. Il n’a jamais été abondant dans les récits coloniaux.
Captivité et dernières observations Le dernier spécimen connu meurt
au zoo de Londres en 1852 après trois ans de captivité. Aucun individu
vivant n’a été observé depuis. Seuls deux spécimens naturalisés sont
conservés dans des collections muséales, notamment à Paris et à Londres
Causes de la disparition La disparition est attribuée à
l’introduction du rat noir (Rattus rattus) et de la mangouste indienne
(Herpestes javanicus), qui ont décimé les populations de rongeurs indigènes
dans les Petites Antilles. La compétition alimentaire, la prédation directe
et la transmission de maladies ont affaibli l’espèce. La colonisation
agricole et la déforestation ont réduit son habitat. L’absence de mesures de
conservation et la faible résilience démographique ont accéléré son
extinction.
Statut actuel et postérité L’espèce est classée comme éteinte par
l’UICN. Elle incarne la fragilité des faunes insulaires face aux
bouleversements coloniaux et écologiques. Elle fait partie du groupe des
Megalomys, tous disparus au XXe siècle, et illustre l’effacement silencieux
des mammifères endémiques des Caraïbes. |
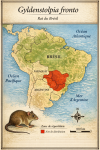
Gyldenstolpia fronto — Rat du Brésil
(moins de 25 cm, mopins de 300 g)

 |
EX
|
Le Gyldenstolpia fronto, ou rat fouisseur géant du Brésil, est
considéré comme éteint depuis le début du XXe siècle, victime de la
fragmentation de son habitat et d’une absence totale de données
contemporaines.
Découverte et répartition initiale L’espèce est décrite en 1888 par
Herluf Winge sous le nom de Kunsia fronto. Elle est ensuite reclassée dans
le genre Gyldenstolpia, qui ne comprend que deux espèces. Le Gyldenstolpia
fronto était présent dans le Chaco humide d’Argentine et dans les savanes
sèches du Brésil, notamment dans le Minas Gerais. Les seuls spécimens connus
proviennent de collectes anciennes, le dernier datant de 1896.
Morphologie et écologie Ce rongeur de grande taille possédait un
crâne robuste, des oreilles arrondies et peu visibles, des yeux petits, une
queue courte et une fourrure dorsale épaisse et hérissée. Il présentait une
morphologie semi-fossoriale, adaptée à la vie souterraine. Son écologie
reste inconnue, mais il est supposé avoir habité les savanes sèches et les
zones herbeuses, avec un mode de vie discret et fouisseur.
Raréfaction et absence de données Aucun spécimen vivant n’a été
observé depuis plus d’un siècle. Des campagnes de recherche ciblées dans le
Chaco argentin et le Cerrado brésilien n’ont donné aucun résultat. L’espèce
est absente des milliers de prélèvements indirects (egagrópilas de rapaces)
réalisés dans ces régions. Aucun fossile récent ni trace génétique n’a été
détecté.
Causes de la disparition La disparition est attribuée à la
fragmentation de son habitat par l’agriculture, l’élevage et l’urbanisation.
L’absence de données contemporaines empêche d’identifier des causes
précises, mais la faible densité, la spécialisation écologique et la
pression anthropique sont les facteurs les plus probables. L’espèce a
disparu sans jamais avoir été étudiée in situ.
Statut actuel et postérité Le Gyldenstolpia fronto est classé comme
éteint par les autorités argentines et comme en danger critique
(possiblement éteint) par l’UICN depuis 2018. Il incarne la disparition
silencieuse des rongeurs sud-américains méconnus, et souligne les lacunes
dans la connaissance et la conservation des petits mammifères fossoriales. |
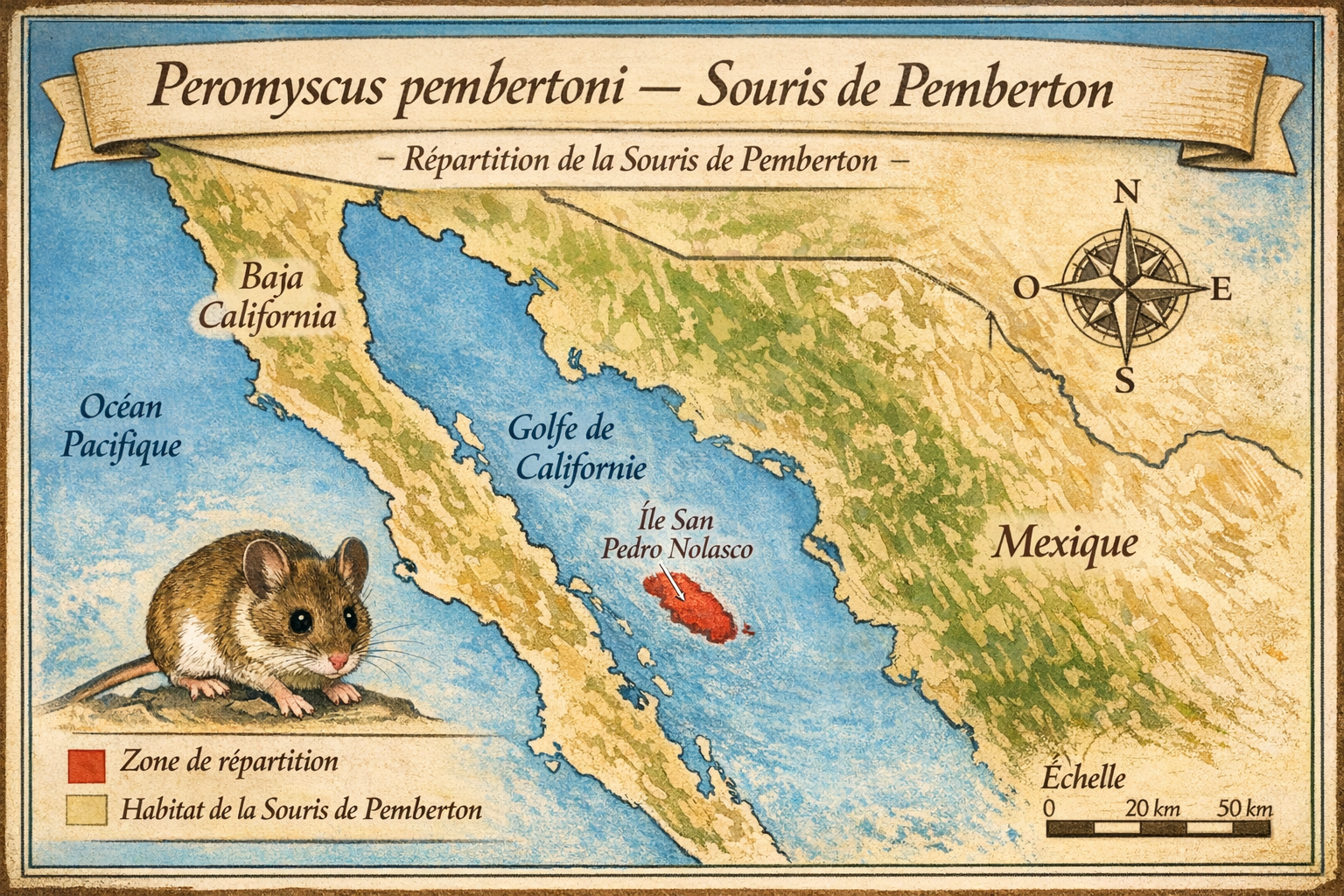
Peromyscus pembertoni — Souris de Pemberton
(moins de 12 cm, moins de 30 g)

 |
EX
|
La souris de Pemberton (Peromyscus pembertoni) a disparu en 1931,
victime de son isolement insulaire, de la compétition interspécifique et de
la fragilité écologique de son habitat.
Découverte et répartition initiale L’espèce est décrite en 1932 par
William H. Burt à partir de spécimens collectés sur l’île San Pedro Nolasco,
dans le golfe de Californie, au large de Sonora (Mexique). Elle était
endémique de cette île aride de 3,7 km², caractérisée par une végétation
clairsemée et une faune limitée. Aucun autre site n’a jamais abrité cette
espèce.
Morphologie et écologie La souris de Pemberton appartenait au
groupe des Peromyscus à pattes blanches. Elle se distinguait par sa taille
modeste, sa fourrure brun-gris, ses grandes oreilles et sa queue velue. Elle
était probablement nocturne, granivore et insectivore, occupant les zones
rocheuses et les crevasses pour se protéger des prédateurs et des
températures extrêmes. Son comportement reste mal connu, faute
d’observations prolongées.
Dernières observations et extinction Le dernier spécimen vivant est
collecté le 26 décembre 1931. Aucune trace n’est relevée lors des
expéditions ultérieures. L’UICN confirme son extinction dans les années
1990. L’espèce n’a jamais été observée sur le continent ni sur d’autres îles
du golfe.
Causes de la disparition La cause principale est la compétition
avec Peromyscus boylii, une espèce continentale introduite accidentellement
sur l’île, probablement par des pêcheurs ou des navigateurs. Ce concurrent
plus adaptable a supplanté la souris de Pemberton dans les niches
écologiques disponibles. L’absence de diversité génétique, la faible
résilience démographique et l’absence de refuge ont accéléré l’extinction.
Aucun programme de conservation n’a été envisagé à l’époque.
Statut actuel et postérité L’espèce est classée comme éteinte par
l’UICN. Elle incarne la vulnérabilité des espèces insulaires face aux
introductions biologiques et à l’absence de surveillance écologique. Elle
est souvent citée comme exemple dans les études sur l’extinction rapide des
petits mammifères insulaires |
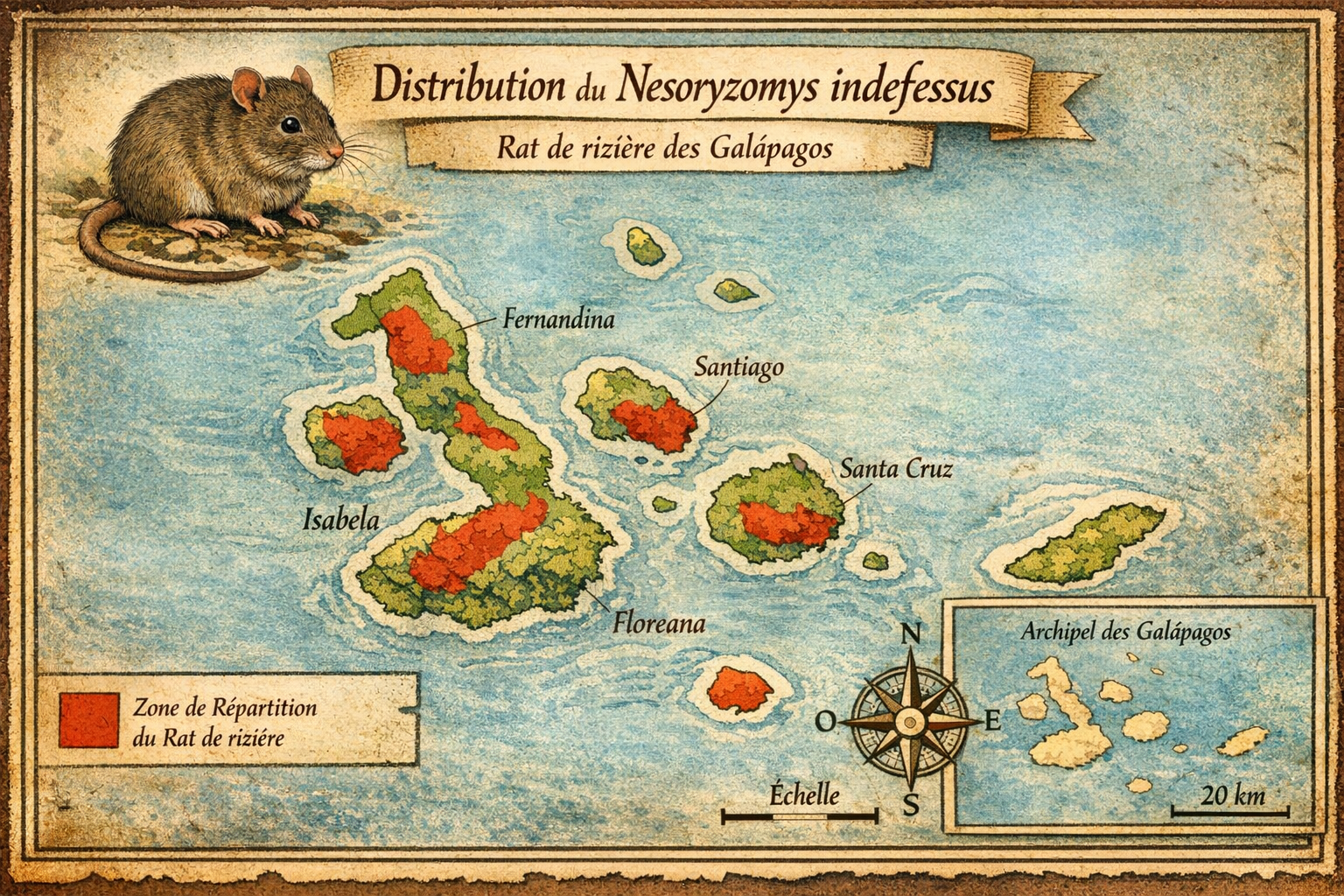
Nesoryzomys indefessus — Rat de rizière des Galapagos
(moins de 18 cm, moins de 80 g)

 |
EX
|
Le Nesoryzomys indefessus, ou rat de rizière des Galápagos, a
disparu dans les années 1930, victime de l’introduction du rat noir et de la
fragilité écologique insulaire.
Découverte et répartition initiale L’espèce est décrite en 1899 par
Oldfield Thomas. Elle était endémique des îles Santa Cruz et Baltra, dans
l’archipel des Galápagos. Elle appartenait au genre Nesoryzomys, un groupe
de rongeurs sigmodontinés spécifiques aux Galápagos, et occupait les zones
boisées et les milieux semi-arides de basse altitude.
Morphologie et écologie Ce rongeur possédait une taille modeste,
une fourrure brun-gris, une queue velue et une dentition adaptée à un régime
omnivore. Il se nourrissait de graines, de fruits, d’insectes et de petits
invertébrés. Son comportement était probablement nocturne et discret, avec
une activité concentrée dans les zones de végétation dense. Il jouait un
rôle écologique important dans la dispersion des graines et le contrôle des
insectes.
Raréfaction et extinction L’espèce devient rare dès le début du XXe
siècle. Les dernières observations confirmées datent des années 1930. Aucune
trace n’a été relevée lors des expéditions scientifiques ultérieures. L’UICN
classe l’espèce comme éteinte, bien que des espèces proches comme
Nesoryzomys narboroughi aient été redécouvertes sur d’autres îles.
Causes de la disparition La principale cause est l’introduction du
rat noir (Rattus rattus) par les navires européens. Ce prédateur
opportuniste a décimé les populations indigènes en détruisant les nids, en
consommant les jeunes et en entrant en compétition pour les ressources
alimentaires. La faible résilience démographique, l’absence de mécanismes de
défense et l’isolement insulaire ont empêché toute adaptation. Aucun
programme de conservation n’a été mis en place à l’époque.
Statut actuel et postérité Le Nesoryzomys indefessus est classé
comme éteint par l’UICN depuis 2019. Il incarne la vulnérabilité des espèces
insulaires face aux introductionsbiologiques et à l’absence de surveillance
écologique. Il est souvent cité comme exemple dans les études sur
l’extinction rapide des petits mammifères insulaires. |
Cétartiodactyles éteints récemment
|
Nom |
Statut |
Parcours et cause
disparition (EX) |
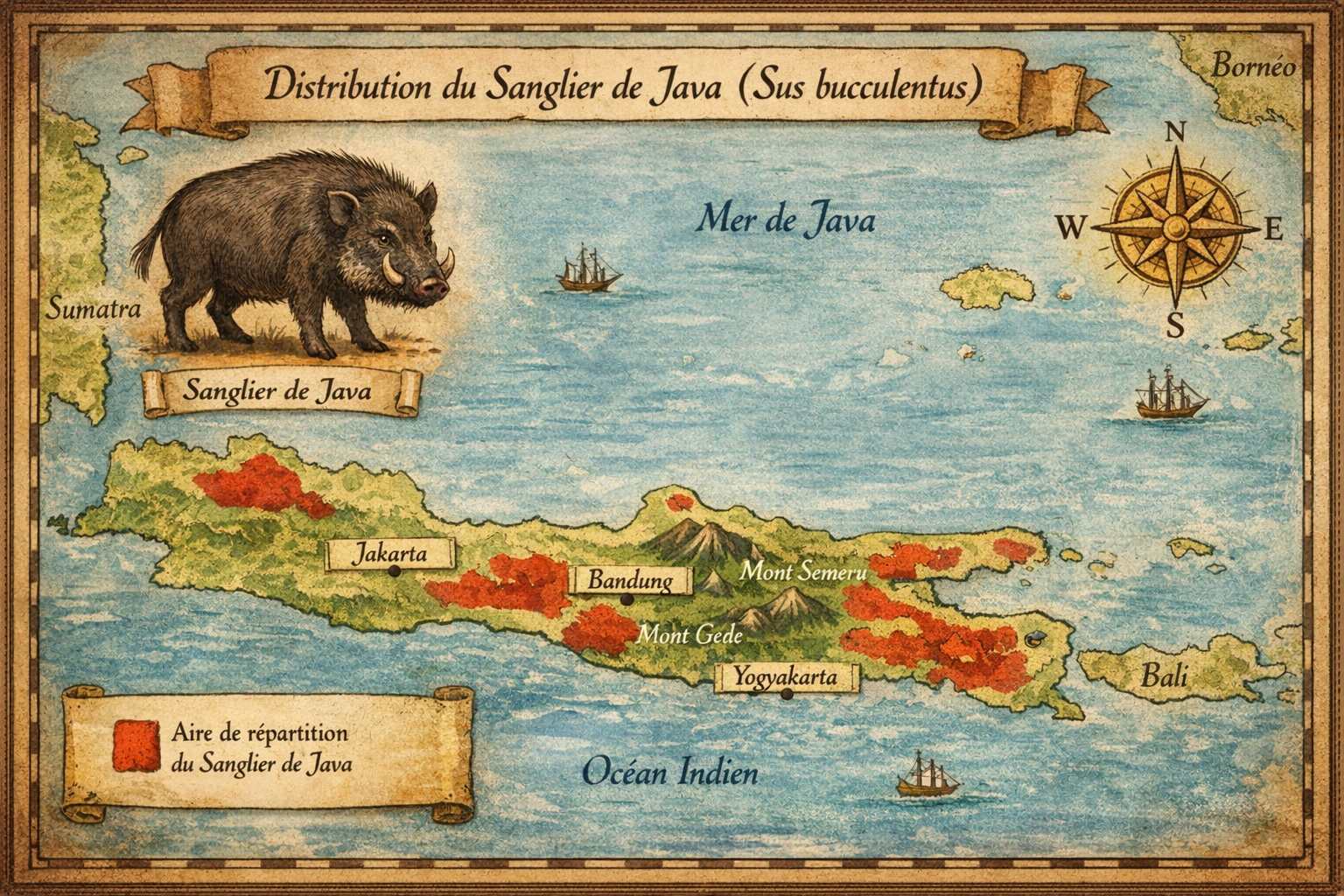
Sanglier
de Java (Sus bucculentus)
(moins de 110 cm, moins de 60 kg)

 |
EX
|
Le sanglier de Java (Sus bucculentus), aussi appelé sanglier du
Vietnam, est considéré comme éteint, bien que son statut taxonomique reste
incertain en raison du faible nombre de spécimens et de données génétiques
disponibles.
Découverte et répartition initiale L’espèce est décrite en 1892 à
partir de deux crânes collectés dans les montagnes du Laos. Un troisième
spécimen est découvert en 1995 dans les monts Annamites. Elle aurait été
endémique des forêts tropicales humides du Laos et du Vietnam, entre 500 et
1 500 mètres d’altitude, dans des zones marécageuses, riveraines et
semi-décidues.
Morphologie et écologie Le sanglier de Java se distinguait par des
joues larges et une dentition robuste, adaptée à une alimentation variée
incluant racines, tubercules, fruits, écorces et graines dures. Il était
légèrement plus grand que le sanglier eurasien (Sus scrofa), avec un poids
estimé entre 80 et 120 kg. Son pelage était probablement dense et sombre.
Les canines bien développées servaient à creuser et à se défendre. Son
comportement était probablement solitaire et nocturne, avec une activité
concentrée dans les zones humides et boisées.
Raréfaction et incertitude taxonomique L’espèce n’a jamais été
observée vivante. Les données sont limitées à quelques crânes et fragments
dentaires. Des analyses génétiques récentes suggèrent qu’il pourrait s’agir
d’une forme locale de Sus scrofa, ce qui rend son statut taxonomique
incertain. L’UICN le considère comme éteint jusqu’à obtention de preuves
plus solides.
Causes de la disparition La disparition est attribuée à la
déforestation, à la fragmentation de l’habitat et à la chasse. Les forêts
des monts Annamites ont été fortement exploitées pour l’agriculture, le bois
et les infrastructures. La chasse locale, motivée par la viande et les
croyances médicinales, a pu décimer les dernières populations. L’absence de
données contemporaines et de programmes de conservation a empêché toute
protection.
Statut actuel et postérité Le sanglier de Java est classé comme
éteint par l’UICN, mais son statut reste sujet à débat. Il incarne les
limites de la taxonomie fondée sur des spécimens fragmentaires et souligne
la nécessité de recherches génétiques et écologiques approfondies dans les
zones tropicales reculées. |
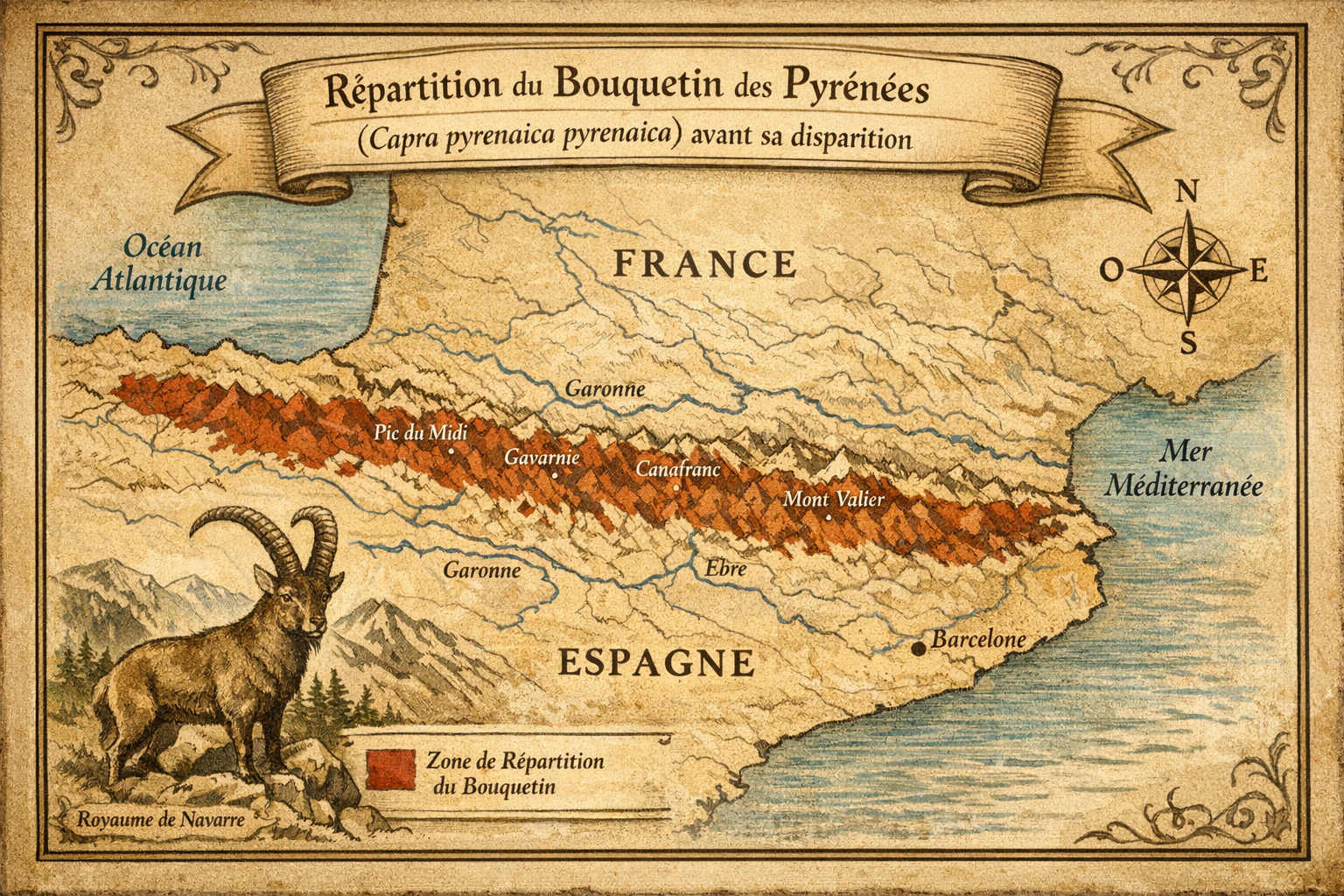
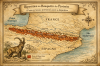
Bouquetin
des Pyrénées (Capra pyrenaica pyrenaica)
(moins de 150 cm, moins de 90 kg)

 |
EX
|
Le Bouquetin des Pyrénées (Capra pyrenaica pyrenaica) a disparu
officiellement en janvier 2000, victime de la chasse intensive, de la
fragmentation de son habitat et de l’absence de mesures de conservation
efficaces.
Répartition et caractéristiques Sous-espèce du bouquetin ibérique,
le Bouquetin des Pyrénées occupait historiquement les versants français et
espagnols de la chaîne pyrénéenne. Il se distinguait par des cornes longues
et incurvées, un pelage sombre marqué de taches noires, et une adaptation
aux milieux escarpés et rocheux. Il vivait en petits groupes dans les zones
d’altitude, entre 1 000 et 2 500 mètres.
Raréfaction et disparition progressive La sous-espèce disparaît du
versant français dès le début du XXe siècle, le dernier individu étant tué
près du lac de Gaube en 1910. En Espagne,
une population relictuelle subsiste dans le parc national d’Ordesa et du
Mont-Perdu, créé en 1918. Malgré cette protection, la population décline
lentement, passant de quelques dizaines à moins de dix individus dans les
années 1990.
Dernier individu et extinction officielle La dernière femelle,
nommée Celia, est retrouvée morte le 6 janvier 2000 dans le parc d’Ordesa,
tuée par la chute d’un arbre. Sa mort
marque l’extinction officielle de la sous-espèce. Des tissus vivants avaient
été prélevés en 1999 pour tenter un clonage, mais les tentatives échouent à
produire une lignée viable.
Causes de la disparition La chasse sportive au XIXe siècle a décimé
les populations, ciblées pour leurs cornes spectaculaires. La fragmentation
de l’habitat par les activités humaines, le dérangement touristique et
l’absence de corridors écologiques ont isolé les derniers groupes. L’absence
de programme de reproduction en captivité ou de translocation a empêché
toute restauration génétique. La faible diversité génétique et la
consanguinité ont aggravé la vulnérabilité démographique.
Statut et postérité Classé comme éteint par l’UICN, le Bouquetin
des Pyrénées est le seul bouquetin ibérique à avoir disparu. Il a été
remplacé par des réintroductions de bouquetins de sous-espèces proches
(Capra pyrenaica victoriae) dans les Pyrénées françaises depuis 2014. Son
extinction est devenue un cas emblématique de perte de biodiversité liée à
la négligence conservatoire. |
Aurochs
(Bos primigenius)
(moins de 180 cm (Garrot), moins de 1 t)

 |
EX
|
L’aurochs (Bos primigenius) a disparu en 1627, victime de la chasse,
de la domestication et de la fragmentation de son habitat forestier.
Origine et répartition historique Apparu il y a environ deux
millions d’années en Inde, l’aurochs migre vers l’Europe, l’Asie et
l’Afrique du Nord. Il devient l’un des grands herbivores du Paléolithique,
représenté dans les grottes de Lascaux, Chauvet et Altamira. Trois
sous-espèces sont identifiées : européenne (Bos primigenius primigenius),
asiatique (Bos primigenius namadicus) et nord-africaine (Bos primigenius
africanus).
Morphologie et écologie L’aurochs mesurait entre 1,60 et 1,80 m au
garrot pour les mâles, avec un poids de 800 à 1 000 kg. Il possédait de
longues cornes en lyre, un pelage brun-noir chez le mâle et brun-rouge chez
la femelle. Il vivait en petits groupes dans les forêts humides, les marais
et les vallées fluviales, se nourrissant de graminées, de glands, d’écorces
et de feuillages.
Domestication et raréfaction Domestiqué vers 8 000 av. J.-C. au
nord du Croissant fertile, il donne naissance aux races bovines actuelles.
Les croisements entre aurochs sauvages et bovins domestiques sont fréquents
jusqu’au Moyen Âge. Dès le XIIIe siècle, l’aurochs ne subsiste plus qu’en
Europe de l’Est, notamment en Pologne, Lituanie et Moldavie.
Extinction et dernier individu Le dernier aurochs connu meurt en
1627 dans la forêt de Jaktorów, en Pologne. Il s’agissait d’une femelle
surveillée par les autorités royales. L’espèce est alors considérée comme
éteinte. Des tentatives de reconstitution ont lieu au XXe siècle, notamment
par les frères Heck, qui créent une race appelée « aurochs de Heck ».
Causes de la disparition La chasse intensive pour la viande et les
trophées a décimé les populations. La domestication a entraîné une pollution
génétique et une réduction des effectifs sauvages. La fragmentation de
l’habitat par l’agriculture et l’urbanisation a limité les zones de refuge.
L’absence de mesures conservatoires et la faible résilience démographique
ont accéléré l’extinction.
Statut et postérité L’aurochs est classé comme éteint par
l’UICN. Il est considéré comme un candidat potentiel pour la désextinction
grâce au séquençage de son ADN ancien. Il reste un symbole de la
domestication et de la disparition des grands herbivores sauvages européens. |
Chiroptères
éteints récemment
|
Nom |
Statut |
Parcours et cause disparition (EX) |
|
Chauve-souris frugivore de Porto Rico (Phyllonycteris
major)
(moins de 12 cm, enverg moins de 45 cm, moins de 60 g)


|
EX
|
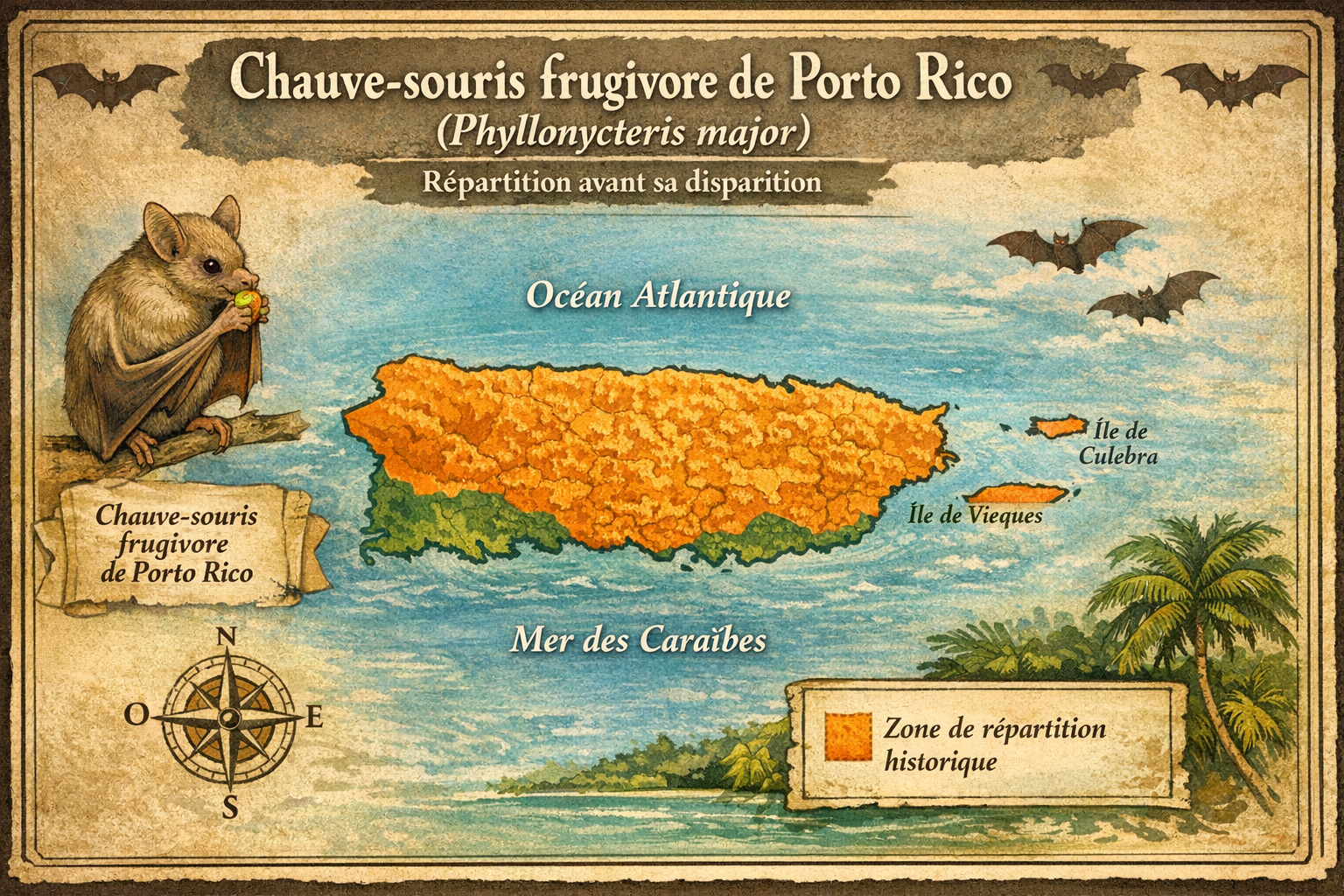
Origine et répartition historique
La Chauve‑souris frugivore de Porto Rico (Phyllonycteris
major) est une espèce éteinte de la famille des
Phyllostomidae elle n’est connue que par des fossiles
découverts à Porto Rico sa présence fossile suggère qu’elle
était autrefois
endémique de Porto Rico,
probablement répandue dans les grottes et forêts de l’île
dans sa description originale de mille neuf cent dix sept,
Harold Elmer Anthony note sa grande proximité avec
Phyllonycteris poeyi, espèce vivante de Cuba, ce qui l’amène
à proposer qu’un ancien lien terrestre ou biogéographique
entre Cuba et Porto Rico pourrait expliquer cette parenté
étroite .
Morphologie
Selon la description fossile, Phyllonycteris major
ressemblait fortement à Phyllonycteris poeyi, mais en plus
grande taille comme les autres membres du genre
Phyllonycteris,
elle possédait probablement un pelage clair, des oreilles
larges, un museau allongé et des adaptations crâniennes
typiques des chauves‑souris nectarivores ou frugivores sa
morphologie suggère un mode de vie similaire à celui des
chauves‑souris frugivores actuelles, avec des ailes adaptées
au vol dans les forêts et les grottes.
Écologie
Bien que l’espèce soit connue uniquement par des fossiles,
son appartenance au genre Phyllonycteris permet d’inférer
une écologie proche de celle des chauves‑souris frugivores
et nectarivores des Caraïbes ces chauves‑souris se
nourrissent généralement de fruits, de nectar et de pollen,
et jouent un rôle important dans la pollinisation et la
dispersion des graines elles fréquentent habituellement les
grottes,
les forêts tropicales, et les zones riches en plantes à
fleurs il est probable que Phyllonycteris major occupait des
habitats similaires à Porto Rico.
Raréfaction et disparition progressive
La disparition de Phyllonycteris major est antérieure aux
temps historiques l’espèce est connue uniquement par des
restes fossiles, ce qui indique une extinction ancienne
plusieurs facteurs probables sont évoqués par les
spécialistes changements climatiques holocènes ayant modifié
la végétation de Porto Rico perturbations écologiques
naturelles dans les grottes possible compétition avec
d’autres chauves‑souris frugivores éventuels impacts humains
très anciens (début de l’occupation précolombienne) la
parenté étroite avec l’espèce cubaine Phyllonycteris poeyi a
conduit Anthony à suggérer que l’espèce aurait pu survivre
plus longtemps que ce que les fossiles indiquent, mais
aucune preuve d’individus vivants n’a jamais été trouvée . |
|
Pipistrelle de Sturdee (Pipistrellus sturdeei)
(moins de 5 cm, enverg moins de 25 cm, moins de 10 g)


|
EX
|
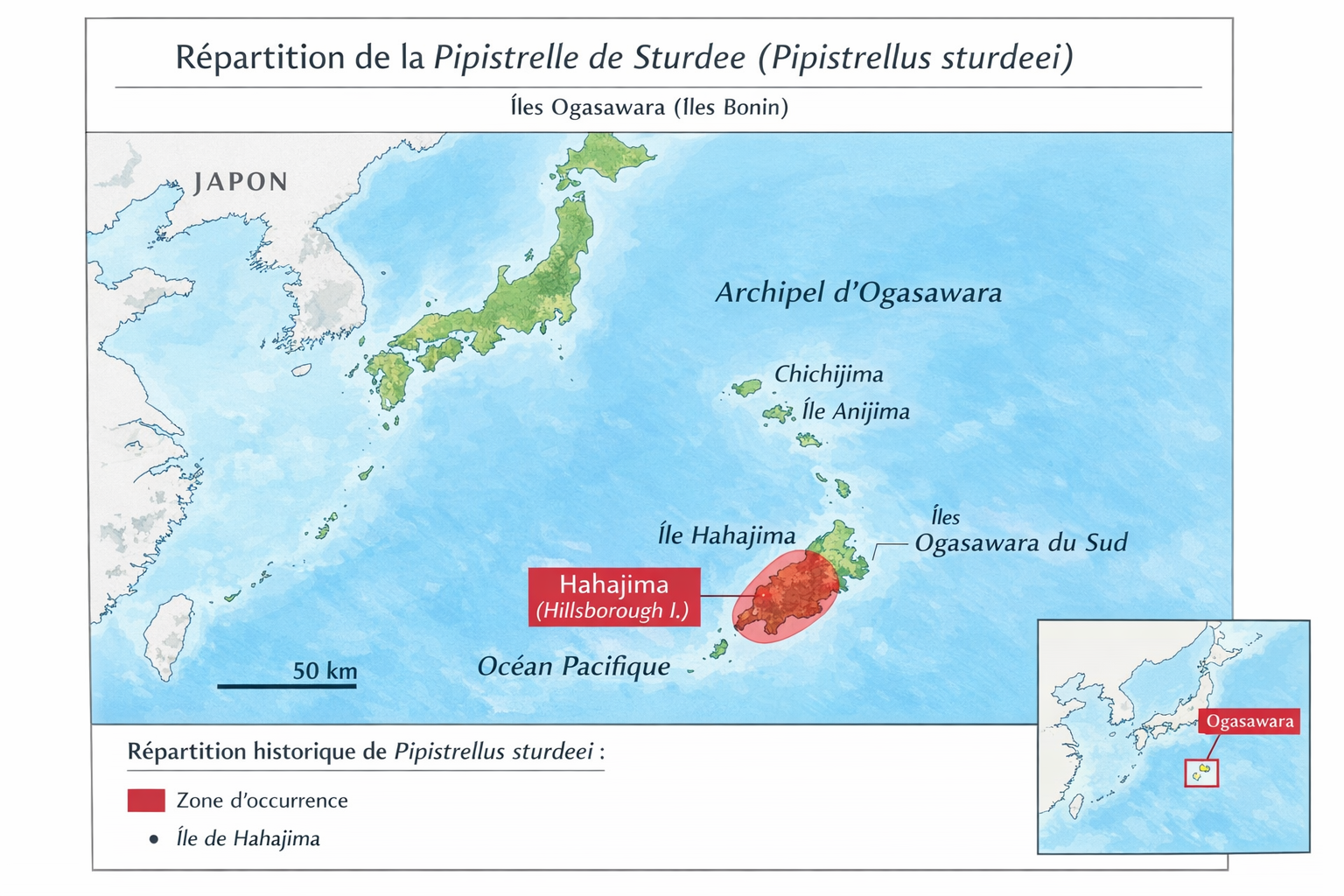
Origine et répartition historique
La Pipistrelle de Sturdee est une espèce décrite en
mille neuf cent quinze par Oldfield Thomas à partir d’un
unique spécimen provenant de l’archipel d’Ogasawara au Japon
aucune autre observation n’a jamais été confirmée certains
auteurs ont suggéré que la localité pourrait être erronée
mais les analyses taxonomiques modernes confirment que
l’étiquette d’origine est correcte et que l’espèce était
bien native des îles Ogasawara elle était donc probablement
endémique de cet archipel isolé du Pacifique occidental.
Morphologie
La description repose uniquement sur l’holotype, une femelle
immature les mesures relevées par Thomas indiquent un corps
et tête de trente sept millimètres, une queue de trente et
un millimètres et un avant‑bras de trente millimètres la
morphologie générale correspond à celle des petites
pipistrelles du genre Pipistrellus, avec un pelage brun, un
museau court, des oreilles modestes et des ailes adaptées au
vol rapide aucune autre donnée morphologique n’existe en
raison de l’absence de spécimens supplémentaires.
Écologie
L’écologie de Pipistrellus sturdeei est inconnue car
l’espèce n’a jamais été observée vivante toutefois, son
appartenance au genre
Pipistrellus permet d’inférer un mode de vie similaire à
celui des pipistrelles actuelles ces chauves‑souris sont
généralement insectivores, chassant en vol à faible altitude
autour des forêts, des lisières et des zones ouvertes les
îles Ogasawara abritent des habitats subtropicaux où
l’espèce aurait probablement exploité les insectes nocturnes
abondants aucune information n’existe sur ses gîtes, mais
les pipistrelles utilisent habituellement les arbres creux,
les fissures rocheuses ou les bâtiments.
Raréfaction et disparition progressive
La Pipistrelle de Sturdee est considérée comme éteinte (EX)
par l’UICN depuis deux mille le dernier et unique spécimen
connu date de
mille huit cent quatre vingt neuf,
ce qui indique une disparition très ancienne malgré des
recherches répétées dans l’archipel, aucune pipistrelle
correspondant à cette espèce n’a jamais été retrouvée les
causes exactes de l’extinction sont inconnues, mais
plusieurs facteurs probables sont évoqués la petite taille
et l’isolement extrême de l’archipel d’Ogasawara la
vulnérabilité aux espèces introduites comme les rats et les
chats la dégradation des habitats liée aux activités
humaines au XIXᵉ siècle la faible résilience démographique
d’une espèce insulaire rare l’UICN note qu’aucune population
n’a été détectée malgré des prospections intensives,
confirmant l’extinction définitive de l’espèce . |
|
Pipistrelle de l’île Christmas (Pipistrellus murray)
(moins de 5 cm, enverg moins de 25 cm, moins de 10 g)


|
EX
|
La Pipistrelle de l’île Christmas était
strictement endémique
de l’île Christmas, un territoire australien de l’océan
Indien. Elle ne vivait
nulle part ailleurs au monde.
Elle était autrefois
commune sur toute l’île,
y compris dans les zones habitées, comme le montrent les
témoignages du début du XXᵉ siècle où les individus
entraient fréquemment dans les maisons . Sa répartition a
commencé à se contracter dans les années 1990, jusqu’à ne
subsister que dans quelques zones forestières avant
l’extinction.
C’était une
très petite chauve‑souris,
la plus petite d’Australie, pesant
3 à 4,5 g.
Son pelage était
brun foncé
avec des
pointes jaunâtres,
ses oreilles étaient
triangulaires,
et son avant‑bras mesurait
30–33 mm.
La longueur tête‑corps atteignait
35–40 mm,
avec une queue de
30–31 mm.
Elle possédait un
uropatagium avec calcar distinct
et une queue dépassant légèrement la membrane .
La Pipistrelle était
nocturne
et se nourrissait d’insectes,
notamment de papillons nocturnes et de coléoptères. Elle
chassait
sous et au‑dessus de la canopée,
souvent le long des lisières forestières. Le jour, elle se
réfugiait dans des
cavités d’arbres,
sous l’écorce
décollée,
dans des
palmes mortes
ou entre un
figuier étrangleur et son hôte.
Elle pouvait se percher seule ou en petites colonies allant
jusqu’à
47 individus.
Les déplacements entre gîtes étaient fréquents, sur des
distances de
14 à 186 m
.
Le déclin de la Pipistrelle de l’île Christmas a été
brutal à partir des années 1990 : encore commune dans les
années 1980, elle avait perdu environ 80 % de sa population
entre 1994 et 2005. En 2009, il ne restait probablement
qu’une vingtaine d’individus, puis un seul, détecté pour la
dernière fois le 26 août 2009, date considérée comme
l’extinction officielle. Les causes identifiées sont presque
entièrement liées aux espèces invasives, en particulier le
serpent‑loup asiatique (Lycodon
capucinus),
responsable estimé de 55 % du déclin, ainsi que le
scolopendre géant et la fourmi folle jaune, qui expliquent
ensemble environ 30 % des pertes. D’autres pressions ont
aggravé la situation : rats noirs, chats, escargot géant
africain et prédation par la crécerelle nankeen. Les études
ont exclu la maladie et démontré une corrélation directe
entre l’expansion du serpent‑loup et l’effondrement de la
population. L’absence d’un programme de captivité mis en
place à temps a été largement critiquée, car l’espèce aurait
probablement pu être sauvée. |
|
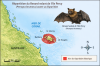
Renard volant de l’île Percy (Pteropus brunneus)
(moins de 25 cm, enverg moins de 110 cm, moins de 1 kg)

|
EX
|
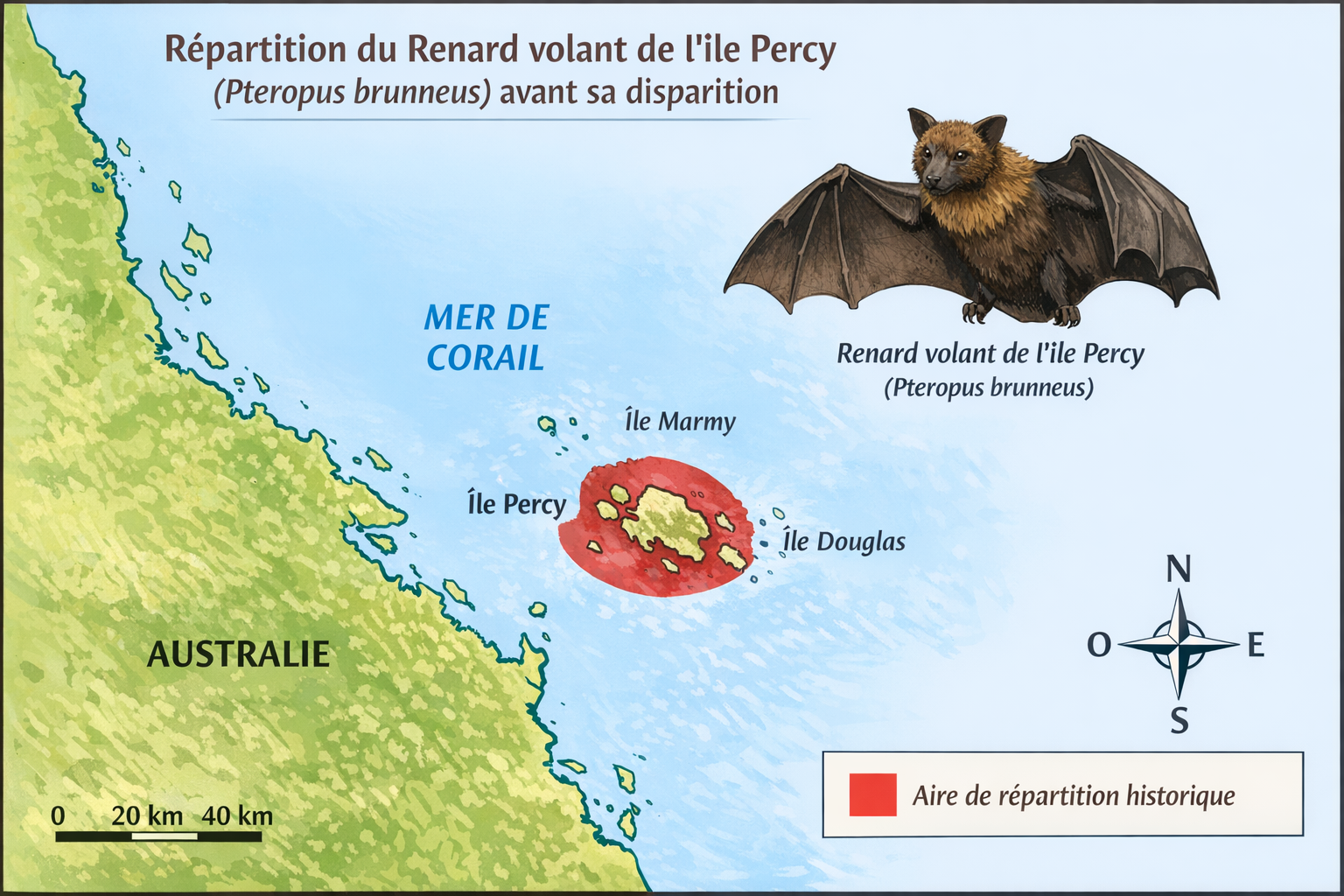
Origine et repartition historique
Le
Renard volant de l’île Percy
(Pteropus
brunneus)
est une espèce de mégachiroptère de la famille des
Pteropodidae,
décrite en 1878 par
George Edward Dobson
à partir d’un unique spécimen collecté en
1874
sur les
îles Percy,
au large du
Queensland,
en
Australie
. Son origine exacte reste incertaine, car certains auteurs
ont suggéré qu’il pourrait s’agir d’un
vagabond
d’une autre espèce mélanésienne, tandis que d’autres travaux
confirment qu’il s’agissait bien d’une espèce distincte,
probablement endémique d’un petit archipel isolé .
Historiquement, l’espèce aurait formé des
colonies importantes
sur les îles Percy et se serait rendue régulièrement sur le
continent australien
pour se nourrir, selon des témoignages du XIXᵉ
siècle . Aucune observation fiable n’a été faite après les
années 1890, malgré de nombreuses
recherches.
Morphologie
Pteropus brunneus
était un
petit renard volant
comparé aux autres espèces australiennes. Le spécimen connu
pesait environ
200 g,
avec une longueur tête‑corps d’environ
210 mm
et un avant‑bras de
118 mm
. Son pelage était
uniformément brun doré,
avec des nuances plus claires sur la tête et la nuque, et
une fourrure légèrement plus longue au niveau du cou. Le
uropatagium
était étroit et partiellement masqué par la fourrure.
L’espèce se distinguait morphologiquement des autres renards
volants australiens, présentant des affinités plus proches
de certaines espèces mélanésiennes, ce qui a alimenté les
débats taxonomiques .
Écologie
Comme les autres membres du genre
Pteropus,
Pteropus brunneus
était probablement
frugivore
et
nectarivore,
se nourrissant de fruits, de fleurs et de pollen. Les
descriptions historiques évoquent des
camps coloniaux
installés dans les forêts des îles Percy, avec des
déplacements réguliers vers le continent pour se nourrir .
L’espèce devait jouer un rôle écologique important dans la
pollinisation
et la
dispersion des graines,
comme la plupart des renards volants. Son habitat supposé
comprenait les
forêts insulaires,
les zones boisées côtières et les milieux tropicaux humides
du Queensland.
Raréfaction et disparition progressive
La disparition de
Pteropus brunneus
est l’une des extinctions les plus rapides et les plus
mystérieuses des mammifères australiens. L’espèce n’a plus
été observée après
1874,
malgré des recherches intensives, et a été déclarée
éteinte
par l’UICN
(catégorie EX) . Plusieurs facteurs ont probablement
contribué à sa disparition. Les
petites populations insulaires
sont extrêmement vulnérables aux perturbations
environnementales, aux cyclones, aux maladies et aux
fluctuations alimentaires. La
déforestation
et les
perturbations humaines
sur les îles Percy au XIXᵉ
siècle ont pu réduire drastiquement son habitat. Les renards
volants sont également sensibles à la
chasse,
aux
incendies
et aux
événements climatiques extrêmes,
qui peuvent décimer des colonies entières. Enfin, la
possibilité que l’espèce ait été déjà
rare
ou
localisée
au moment de sa découverte a pu accélérer son extinction.
Les derniers témoignages du XIXᵉ
siècle évoquent encore des colonies nombreuses, mais
celles‑ci semblent avoir disparu en quelques décennies
seulement, faisant de
Pteropus brunneus
un exemple emblématique d’extinction rapide d’un mammifère
insulaire . |
|
Grand renard volant des Palaos (Pteropus pilosus)
(moins de 25 cm, enverg moins de 120 cm, moins de 1 kg)


|
EX
|
Origine et repartition historique
Le
Grand renard volant des Palaos
(Pteropus
pilosus)
était un mégachiroptère endémique des
îles Palaos,
en
Micronésie,
où il ne vivait nulle part ailleurs. L’espèce n’est connue
que par
deux spécimens
collectés avant
1874,
ce qui indique qu’elle était déjà extrêmement rare au moment
de sa découverte. Aucun témoignage ne mentionne sa présence
en dehors de l’archipel, et sa répartition historique semble
avoir été strictement limitée aux forêts insulaires des
Palaos. L’UICN confirme que l’espèce était
endémique
et n’a jamais été observée ailleurs dans le Pacifique.
Morphologie
Pteropus pilosus
était un renard volant de
taille moyenne,
avec une envergure d’environ
60 cm.
Son pelage était
brunâtre,
avec des
poils argentés
sur le ventre, ce qui lui donnait un aspect contrasté. Sa
morphologie générale était similaire à celle des autres
espèces du genre
Pteropus,
mais les spécimens montrent une fourrure ventrale
particulièrement longue et brillante. L’espèce a été décrite
scientifiquement en
1908
par
Knud Andersen,
à partir des deux spécimens collectés avant 1874.
Ecologie
Aucune donnée directe n’existe sur l’écologie de
Pteropus pilosus,
car l’espèce avait déjà disparu avant que des observations
naturalistes détaillées puissent être réalisées. L’UICN
précise qu’aucune
information écologique fiable
n’a été enregistrée. Cependant, par analogie avec les autres
renards volants du Pacifique, il est probable que l’espèce
ait été
frugivore
et
nectarivore,
se nourrissant de fruits, de fleurs et de pollen, et jouant
un rôle important dans la
pollinisation
et la
dispersion des graines
dans les forêts insulaires. Comme les autres
Pteropodidae,
elle devait former de petites colonies perchées dans les
arbres et se déplacer la nuit pour se nourrir.
Raréfaction et disparition progressive
Le
Grand renard volant des Palaos
est considéré comme
éteint
depuis
1874,
et l’UICN le classe dans la catégorie
EX – Extinct.
Les causes exactes de sa disparition restent inconnues, mais
plusieurs facteurs probables sont évoqués. La
chasse excessive
par les populations locales est considérée comme une cause
majeure, comme le suggère la notice de
Wikipedia.
Les renards volants sont traditionnellement chassés pour
leur viande dans de nombreuses îles du Pacifique, et une
espèce insulaire à faible effectif est particulièrement
vulnérable. La
faible taille de la population,
le
caractère insulaire,
les
événements climatiques extrêmes
(typhons), la
déforestation locale
et l’absence de diversité génétique ont probablement
accéléré le déclin. L’espèce n’a jamais été revue malgré des
recherches répétées, ce qui confirme une extinction très
rapide, probablement survenue en quelques décennies
seulement. L’UICN note que l’espèce n’a
jamais été observée depuis 1874,
malgré des inventaires intensifs. |
|
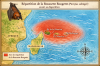
Roussette rougette (Pteropus subniger)
(moins de 20 cm, enverg moins de 90 cm, moins de 500 g)

|
EX
|
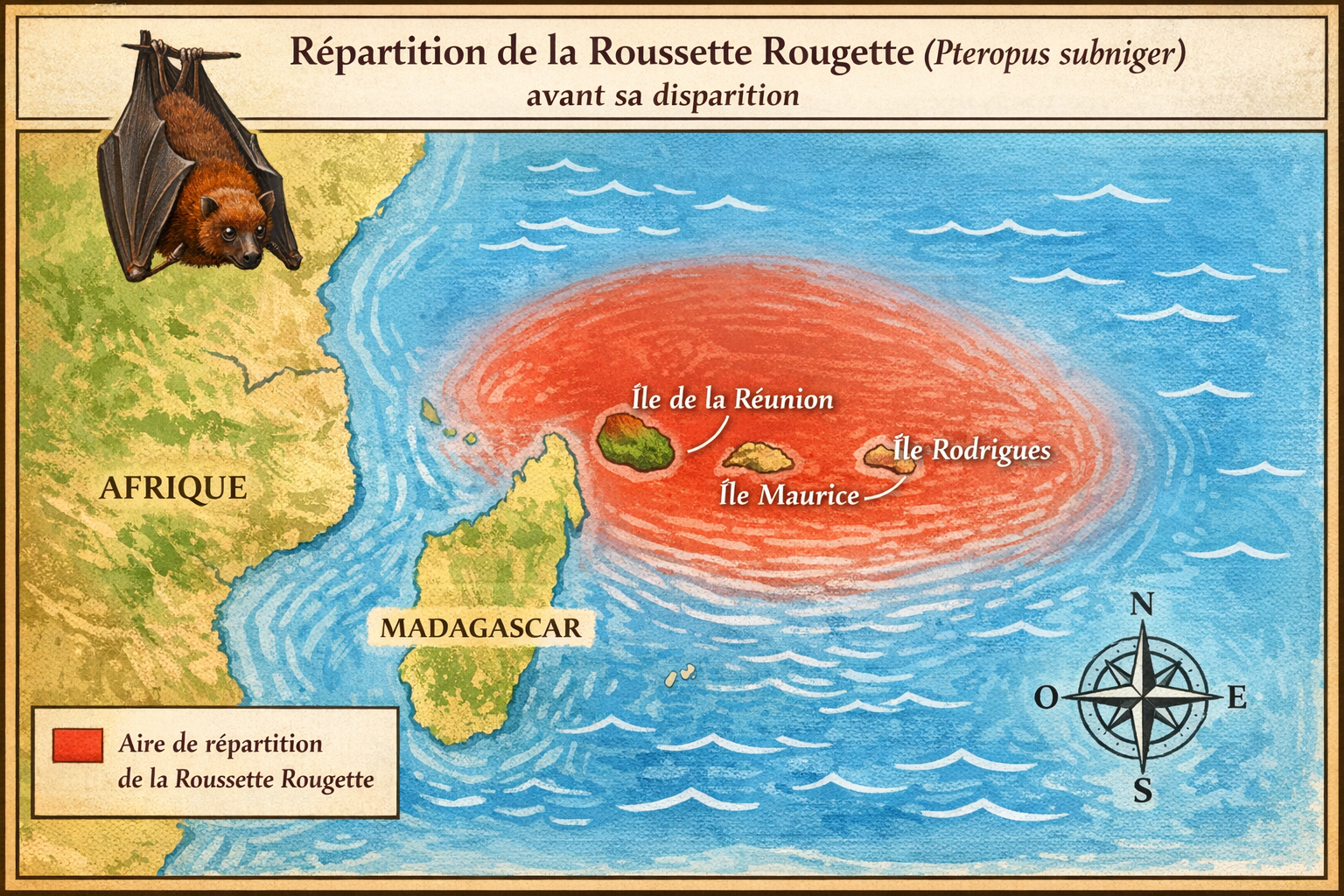
Origine et repartition historique
La
Roussette rougette
(Pteropus
subniger),
également appelée
Small Mauritian flying fox
ou
Dark flying fox,
était un renard volant endémique des
Mascareignes,
où elle ne vivait que sur
Maurice
et
La Réunion.
Les premiers spécimens ont été décrits en
1792
par
Robert Kerr.
L’espèce était autrefois
abondante,
occupant les forêts humides, les zones montagneuses et les
grands arbres creux des deux îles. Elle formait des colonies
pouvant atteindre
plus de 400 individus,
ce qui témoigne d’une présence autrefois massive dans les
paysages forestiers mauriciens et réunionnais. Sa
répartition historique était donc strictement limitée à ces
deux îles volcaniques de l’océan Indien, sans aucune
présence ailleurs.
Morphologie
Pteropus subniger
était une chauve‑souris de
taille moyenne,
avec un avant‑bras mesurant environ
95 à 99 mm
et une fourrure
longue, dense et soyeuse.
Sa coloration était
brun noirâtre,
avec une tête
grisonnante
et un
collier brun orangé
très caractéristique. Le museau était long et effilé, les
yeux grands, et les oreilles petites et dissimulées dans la
fourrure. L’espèce ne possédait
pas de queue,
et son uropatagium se réduisait à une fine membrane interne.
Les dents étaient
petites et délicates,
ce qui suggère un régime alimentaire basé sur le nectar et
les fruits mous. Ces traits morphologiques la rapprochent du
groupe d’espèces de
Pteropus hypomelanus,
selon la classification d’Andersen.
Écologie
La Roussette rougette était
nocturne
et se réfugiait dans les
grands arbres creux,
parfois dans des grottes, formant des colonies importantes.
Les observations historiques indiquent qu’elle quittait ses
gîtes au crépuscule et revenait avant l’aube. Son régime
alimentaire était probablement
nectarivore
et
frugivore,
ce que suggèrent ses dents fines et les comportements
rapportés. Elle jouait un rôle écologique essentiel dans la
pollinisation
et la
dispersion des graines
des forêts mauriciennes et réunionnaises. Les témoignages du
XVIIIᵉ
siècle décrivent une espèce autrefois très commune, présente
même dans les zones habitées, ce qui montre qu’elle
supportait une certaine proximité avec l’activité humaine
avant son déclin.
Raréfaction et disparition progressive
La disparition de
Pteropus subniger
est attribuée à une combinaison de facteurs humains et
environnementaux. L’espèce a été
massivement chassée
pour sa viande et sa graisse, très recherchées par les
colons européens et les populations locales, comme le
rapportent les récits du XVIIIᵉ
siècle. Les « bat‑trees »,
arbres creux abritant des colonies, étaient activement
exploités, parfois fumés pour capturer ou tuer les animaux.
La
déforestation
rapide des Mascareignes, liée à l’agriculture coloniale, a
détruit une grande partie de ses habitats de gîte et
d’alimentation. Les populations se sont effondrées au XIXᵉ
siècle : l’espèce a disparu de
La Réunion
vers
1862,
puis de
Maurice
entre
1864
et 1873,
selon l’UICN,
qui la classe aujourd’hui comme
éteinte (EX).
Aucun individu n’a été observé depuis le milieu du XIXᵉ
siècle. La combinaison de la chasse intensive, de la perte
d’habitat, de la faible résilience des populations
insulaires et de la fragmentation écologique a conduit à une
extinction rapide et irréversible. |
Périssodactyles éteints récemment
|
Nom |
Statut |
Parcours et cause disparition (EX) |
|
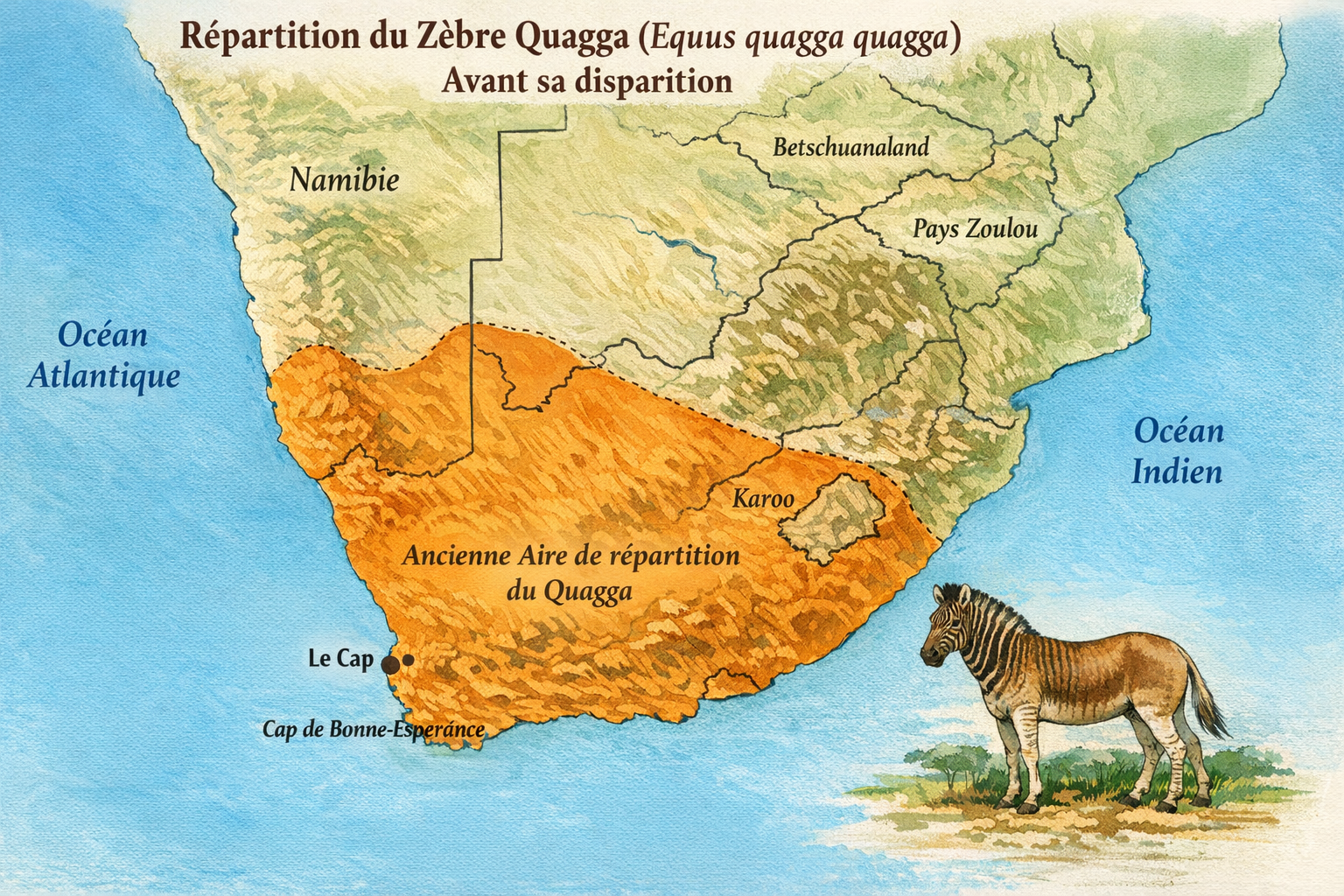
Zèbre Quagga (Equus quagga quagga)


|
EX
|
Le
Quagga
(Equus
quagga quagga)
était une sous‑espèce méridionale du
zèbre des plaines,
endémique d’Afrique australe. Il occupait autrefois les
prairies arides,
les
savannes ouvertes
et les
bushvelds
du
Sud de l’Afrique,
principalement en
Afrique du Sud,
dans les régions du
Cap,
du
Karoo,
de l’État
libre d’Orange
et du
Transvaal.
Sa répartition historique était limitée aux zones les plus
sèches et herbeuses, où il vivait en grands troupeaux mixtes
avec d’autres herbivores. Les analyses génétiques modernes
ont confirmé que le Quagga n’était pas une espèce distincte,
mais une
sous‑espèce locale
du zèbre des plaines, adaptée aux environnements plus arides
du sud du continent.
Le Quagga se distinguait par une apparence unique parmi les
zèbres. L’avant du corps était
fortement rayé,
comme chez les autres zèbres, mais les rayures s’estompaient
progressivement vers l’arrière pour laisser place à une
robe brun‑roux unie
sur la croupe et les pattes. Cette transition spectaculaire
lui donnait un aspect « mi‑zèbre, mi‑cheval ». Il mesurait
environ
1,25 m à 1,35 m
au garrot, avec une morphologie robuste adaptée à la course
en milieu ouvert. Sa crinière était courte et dressée, et
son museau sombre contrastait avec les rayures faciales. Les
spécimens naturalisés montrent une grande variabilité
individuelle, ce qui reflète probablement une diversité
phénotypique importante dans les populations sauvages.
Le Quagga était un
herbivore strict,
spécialisé dans la consommation d’herbes
dures
des savannes sèches. Il vivait en
troupeaux sociaux,
souvent associés aux
gnous bleus
et aux
autres zèbres,
formant des communautés mixtes qui amélioraient la vigilance
contre les prédateurs. Comme les autres zèbres, il jouait un
rôle écologique essentiel dans le
pâturage,
la
dispersion des graines
et la
structuration des prairies.
Il se déplaçait en fonction des ressources en eau et en
herbe, occupant des territoires saisonniers dans les plaines
sud‑africaines. Les récits du XIXᵉ siècle décrivent des
troupeaux très nombreux, parfois de plusieurs centaines
d’individus.
La disparition du Quagga est directement liée à l’expansion
coloniale européenne en Afrique australe. À partir du début
du
XIXᵉ siècle,
l’espèce a été
massivement chassée
pour sa viande, sa peau et pour protéger les pâturages
destinés au bétail. Les colons considéraient le Quagga comme
un
concurrent des moutons et des bovins,
ce qui entraîna une extermination systématique. La
dégradation des habitats,
la
fragmentation des troupeaux
et l’absence de mesures de conservation ont accéléré son
déclin. Dès les années
1850,
il avait disparu de la plupart de son aire naturelle. Le
dernier Quagga sauvage aurait été tué vers
1878
dans l’État libre d’Orange. Le dernier individu captif est
mort au
zoo d’Amsterdam
le
12 août 1883,
marquant l’extinction officielle de la sous‑espèce.
Au XXᵉ siècle, des analyses génétiques ont permis de
confirmer son statut de sous‑espèce, ce qui a conduit au
lancement du
Quagga Project,
un programme visant à sélectionner des zèbres des plaines
présentant des caractéristiques proches du Quagga afin de
reconstituer un phénotype similaire. Bien que ces animaux ne
soient pas des Quaggas au sens strict, ils témoignent de
l’intérêt scientifique et culturel pour cette sous‑espèce
disparue. |
|
Rhinocéros noir d’Afrique de l’Ouest (Diceros bicornis
longipes)

 |
EX
|
Le
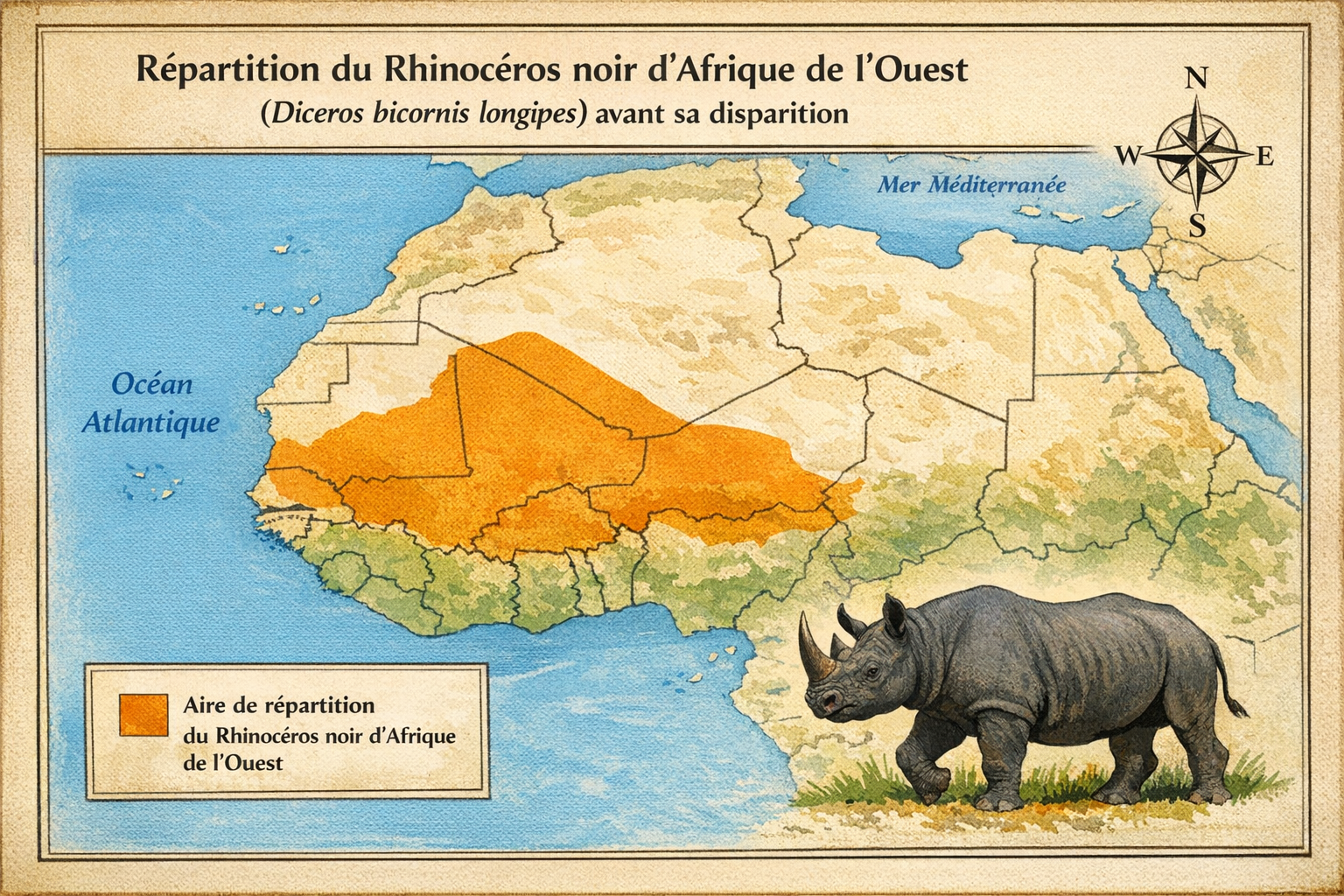
Rhinocéros noir d’Afrique de l’Ouest
(Diceros
bicornis longipes)
était une sous‑espèce du rhinocéros noir, endémique de l’Afrique
de l’Ouest et du centre‑ouest.
Il occupait autrefois un vaste territoire s’étendant du
Tchad
au
Cameroun,
en passant par la
République centrafricaine,
le
nord du Nigeria,
le
nord du Bénin
et parfois le
nord du Mali.
Sa répartition suivait les zones de
savane arborée,
de
brousse sèche
et de
forêts claires soudaniennes,
où il trouvait des arbustes épineux adaptés à son régime
alimentaire. Cette sous‑espèce représentait la limite
occidentale de la distribution du rhinocéros noir sur le
continent africain.
Comme les autres rhinocéros noirs,
Diceros bicornis longipes
possédait deux cornes, dont la première pouvait atteindre
1 mètre.
Il mesurait environ
1,40 à 1,60 m
au garrot et pesait entre
800 et 1 300 kg.
Son corps était massif, avec une peau gris foncé et un
museau préhensile en forme de
bec crochu,
adapté pour saisir les rameaux et les feuilles. Les
descriptions indiquent qu’il était légèrement plus
élancé
que les sous‑espèces orientales, avec des membres
proportionnellement plus longs, ce qui correspond à son
adaptation aux savanes plus ouvertes de l’Afrique de l’Ouest
Le rhinocéros noir d’Afrique de l’Ouest était un
nourrisseur sélectif
se nourrissant principalement de
feuilles,
rameaux,
arbustes épineux
et
plantes ligneuses.
Il vivait en
solitaire,
sauf pour les femelles accompagnées de leurs petits. Son
territoire dépendait de la disponibilité en eau, qu’il
devait trouver régulièrement, bien qu’il puisse survivre
dans des zones relativement sèches. Il jouait un rôle
écologique essentiel dans la
régénération de la végétation,
en contrôlant la croissance des arbustes et en maintenant
l’équilibre des savanes. Ses prédateurs naturels étaient
rares, les jeunes pouvant occasionnellement être victimes de
lions ou de hyènes, mais l’espèce était surtout vulnérable
aux activités humaines.
La disparition de
Diceros bicornis longipes
est directement liée à l’intensification du
braconnage
pour la corne de rhinocéros, dont la valeur a explosé au XXᵉ
siècle. Dès les années
1970,
la sous‑espèce avait perdu plus de
90 %
de sa population. Les conflits armés dans la région du
Sahel,
la
dégradation des habitats,
la
raréfaction des points d’eau
et l’absence de mesures de conservation efficaces ont
accéléré son déclin. Dans les années
1980,
il ne subsistait plus que quelques dizaines d’individus,
principalement dans le
nord du Cameroun.
Les derniers individus confirmés ont été observés dans le
Parc national de la Bénoué
et le
Parc national de Bouba‑Ndjida,
mais malgré des recherches intensives, aucun rhinocéros n’a
été détecté après
2006.
En
2011,
l’UICN
a officiellement déclaré la sous‑espèce
éteinte,
faisant du rhinocéros noir d’Afrique de l’Ouest l’un des
exemples les plus emblématiques d’extinction récente d’un
grand mammifère africain. Sa disparition résulte d’une
combinaison de
braconnage incontrôlé,
d’instabilité
politique,
d’absence
de protection réelle
et de la
faible résilience
des populations isolées. |
|
Cheval de Tarpan (Equus ferus ferus)

 |
EX
|
Le
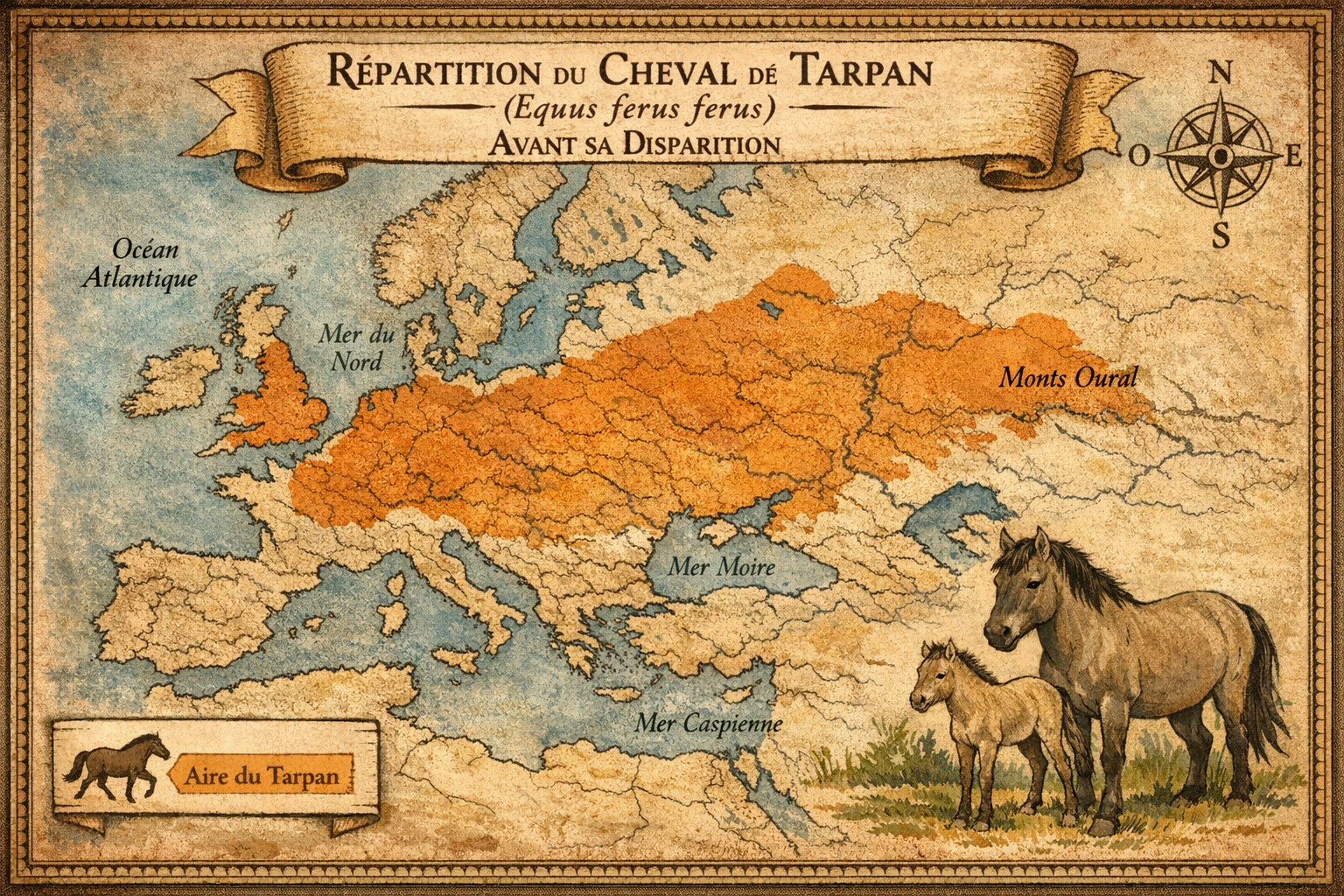
Tarpan
était le cheval sauvage d’Europe
orientale
et des
steppes pontiques,
considéré comme la forme occidentale du cheval sauvage
eurasiatique. Il descendait probablement d’un ancêtre commun
aux chevaux domestiques et aux chevaux de Przewalski, mais
représentait une lignée distincte adaptée aux
steppes froides,
aux
forêts claires
et aux
prairies ouvertes.
Sa répartition historique s’étendait de la
Pologne
et de la
Lituanie
jusqu’à l’Ukraine,
le
sud de la Russie,
le
Kazakhstan occidental
et les régions steppiques proches de la mer Noire. Il
existait deux formes écologiques : le
Tarpan des steppes,
plus clair et plus élancé, et le
Tarpan forestier,
plus sombre et plus massif, vivant dans les forêts d’Europe
centrale et orientale. Les populations étaient autrefois
très répandues, mais déjà fragmentées au Moyen Âge.
Le Tarpan était un cheval de
petite taille,
mesurant environ
1,30 à 1,40 m
au garrot. Sa robe était généralement
gris souris,
parfois tirant vers le
brun‑dun,
avec une
raie de mulet
dorsale et des
zébrures
légères sur les membres, caractéristiques des chevaux
primitifs. Sa crinière était
courte,
souvent
dressée,
et sa queue relativement fournie. Il possédait une tête
robuste, un front large et un corps compact, adapté aux
hivers rigoureux des steppes. Les descriptions historiques
évoquent un animal vif, méfiant, difficile à approcher, avec
une morphologie proche de celle des chevaux primitifs
représentés dans l’art rupestre paléolithique.
Le Tarpan était un
herbivore
spécialisé dans les
graminées
et les
plantes steppiques,
capable de survivre dans des milieux pauvres et des
conditions climatiques extrêmes. Il vivait en
petits groupes familiaux,
menés par une jument dominante, et se déplaçait selon la
disponibilité des pâturages. Il jouait un rôle écologique
important dans le
maintien des prairies,
en empêchant l’embroussaillement et en favorisant la
diversité végétale. Comme les autres équidés sauvages, il
était migrateur à petite échelle, alternant entre zones
forestières et steppiques selon les saisons. Ses prédateurs
naturels incluaient le
loup
et, plus anciennement, le
lynx
et l’ours
brun,
mais sa principale menace devint l’activité humaine.
La disparition du Tarpan résulte d’une combinaison de
chasse,
de
domestication forcée,
de
hybridation,
de
destruction des habitats
et de
conflits avec l’élevage.
Dès le Moyen Âge, il fut considéré comme un
nuisible
par les paysans, car il concurrençait les chevaux
domestiques et endommageait les cultures. Les populations
forestières d’Europe centrale furent éliminées dès le
XVIIᵉ siècle.
Les derniers Tarpans des steppes survécurent en
Ukraine
et dans le
sud de la Russie,
mais furent intensément chassés au XVIIIᵉ et XIXᵉ siècles.
Le dernier individu sauvage connu aurait été tué en
1879
dans la région de
Kherson.
Un dernier Tarpan captif, probablement issu d’une population
semi‑sauvage, mourut en
1887
dans un domaine privé en
Pologne,
marquant l’extinction officielle de la sous‑espèce.
Au XXᵉ siècle, des programmes comme le
Konik polski
ou le
Tarpan reconstitué
ont tenté de recréer un phénotype proche du Tarpan à partir
de chevaux rustiques, mais ces animaux ne sont pas des
Tarpans véritables, seulement des
reconstitutions morphologiques. |
Soricomorphes éteints récemment
|
Nom |
Statut |
Parcours et cause disparition (EX) |
|
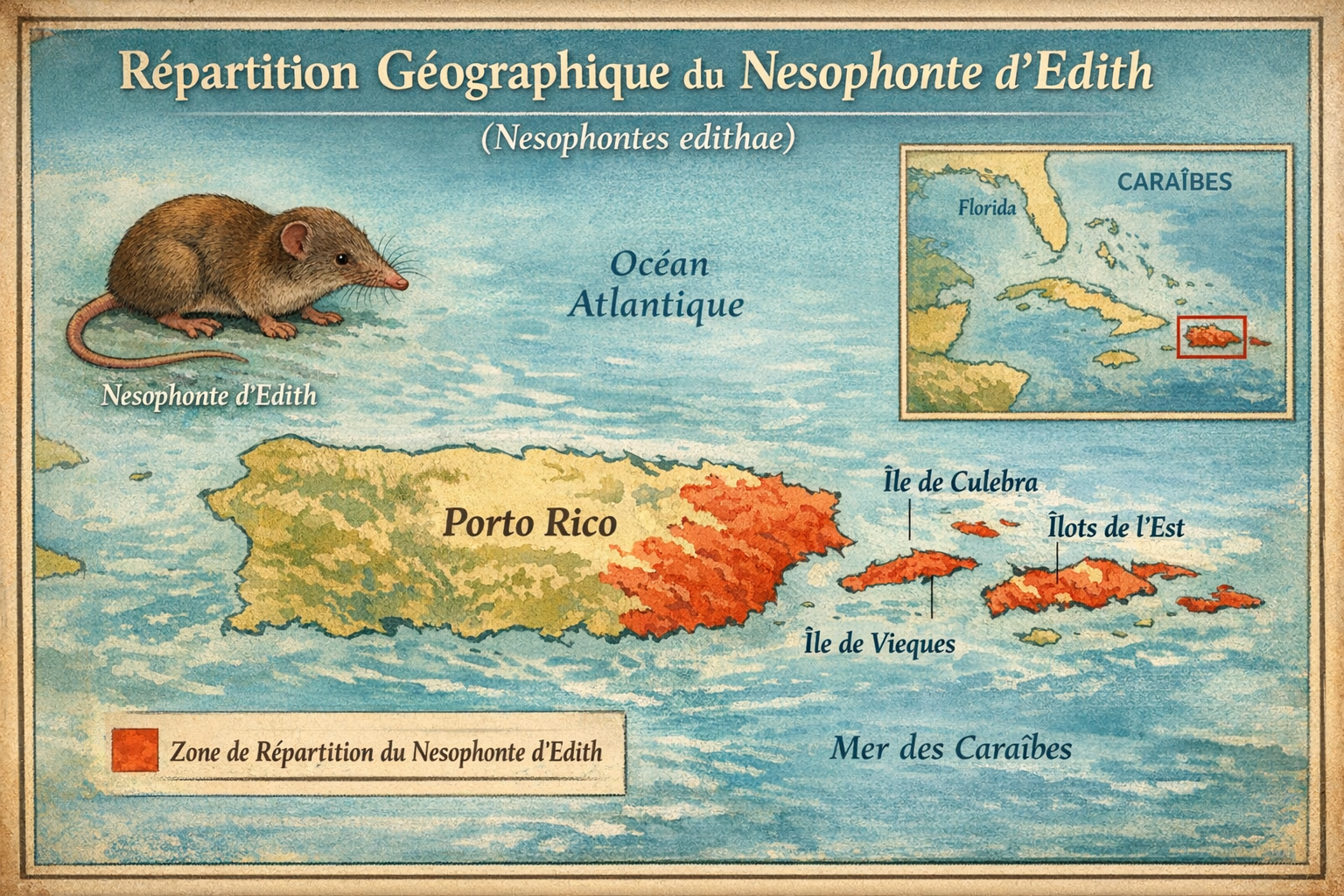
Nesophonte d’Edith (Nesophontes edithae)

 |
EX
|
Le
Nesophonte d’Edith
(Nesophontes
edithae)
appartenait à la famille éteinte des
Nesophontidae,
un groupe de petits insectivores endémiques des
Antilles.
Cette espèce était strictement
endémique de l’île de Saint‑John,
dans les
Îles Vierges américaines,
où elle occupait autrefois les forêts sèches, les zones
broussailleuses et les milieux rocheux. Les fossiles
indiquent qu’elle était présente sur l’île depuis plusieurs
milliers d’années, probablement depuis le
Pléistocène tardif,
et qu’elle représentait l’un des derniers survivants d’un
groupe autrefois plus diversifié dans les Grandes et Petites
Antilles. Sa répartition historique semble avoir été très
restreinte, limitée à Saint‑John, sans preuve de présence
sur les îles voisines comme Saint‑Thomas ou Tortola.
Nesophontes edithae
était un petit mammifère insectivore, de taille comparable à
une
musaraigne,
avec un corps mesurant environ
10 à 12 cm
et un poids estimé entre
20 et 40 g.
Il possédait un
museau allongé,
des
dents pointues
adaptées à un régime insectivore, et un crâne relativement
robuste pour sa taille. Les analyses morphologiques montrent
qu’il avait des membres courts, une queue modérément longue
et une fourrure probablement dense. Ses dents tricuspidées
et son museau étroit rappellent les
Soricidae,
mais les Nesophontidae formaient une lignée totalement
distincte, endémique des Caraïbes depuis plusieurs millions
d’années.
L’écologie du Nesophonte d’Edith est déduite de ses proches
parents et de ses caractéristiques dentaires. Il était très
probablement
insectivore,
se nourrissant d’insectes, d’arthropodes, de larves et
peut‑être de petits invertébrés du sol. Il vivait dans la
litière forestière,
sous les rochers, dans les crevasses et les zones humides
des forêts sèches tropicales. Comme les autres Nesophontes,
il devait être
nocturne
ou
crépusculaire,
actif au sol, et jouait un rôle écologique important dans le
contrôle des populations d’insectes. Les fossiles montrent
qu’il cohabitait autrefois avec des oiseaux endémiques, des
reptiles et d’autres petits mammifères aujourd’hui disparus
ou introduits.
La disparition de
Nesophontes edithae
est étroitement liée à l’arrivée des
Européens
dans les Caraïbes et à l’introduction de
prédateurs exotiques,
en particulier la
ratte noire
(Rattus
rattus)
et la
ratte brune
(Rattus
norvegicus).
Ces rongeurs, arrivés dès le XVIᵉ siècle, ont provoqué une
compétition alimentaire,
une
prédation directe
et une
transmission de maladies
auxquelles les Nesophontes n’étaient pas adaptés. La petite
taille de l’île de Saint‑John a rendu l’espèce
particulièrement vulnérable. La
déforestation,
les
incendies,
la
modification des habitats
et l’expansion des plantations ont aggravé la situation. Les
études paléontologiques montrent que l’espèce a survécu
jusqu’à une période très récente, probablement jusqu’au
XIXᵉ siècle,
voire au début du
XXᵉ siècle,
avant de disparaître complètement. L’UICN la classe
aujourd’hui comme
éteinte (EX).
Sa disparition fait partie d’un effondrement plus large des
petits mammifères endémiques des Antilles, dont plusieurs
espèces de
Nesophontes
ont disparu en quelques siècles seulement. |
|
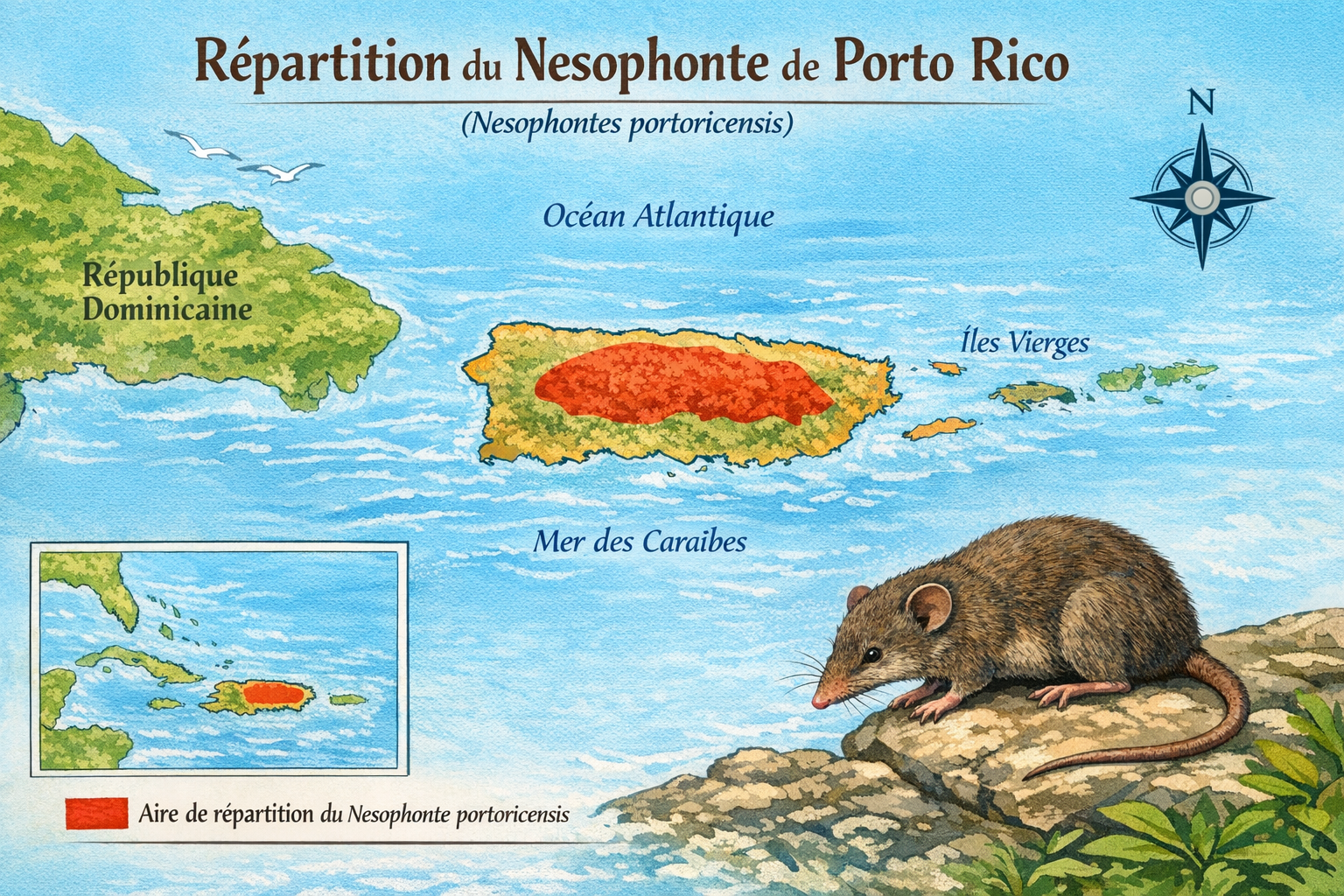
Nesophonte de Porto Rico (Nesophontes portoricensis)

 |
EX
|
Le
Nesophonte de Porto Rico
(Nesophontes
portoricensis)
appartenait à la famille éteinte des
Nesophontidae,
un groupe de petits insectivores endémiques des
Antilles.
Cette espèce était strictement
endémique de Porto Rico,
où elle occupait autrefois les forêts humides, les forêts
sèches, les zones broussailleuses et les milieux rocheux de
l’île. Les fossiles indiquent qu’elle était présente depuis
le
Pléistocène tardif,
et qu’elle cohabitait avec d’autres espèces endémiques
aujourd’hui disparues. Sa répartition historique semble
avoir couvert l’ensemble de l’île, mais elle était
probablement plus abondante dans les zones de forêts denses
et de litière profonde.
Aucun fossile n’indique une présence sur les îles voisines
comme Vieques ou Culebra, ce qui confirme son endémisme
strict à Porto Rico.Nesophontes
portoricensis
était un petit mammifère insectivore, de taille comparable à
une
musaraigne,
avec un corps d’environ
10 à 12 cm
et un poids estimé entre
20 et 40 g.
Il possédait un
museau long et étroit,
des
dents fines et pointues
adaptées à la capture d’insectes, et un crâne robuste pour
sa taille. Ses dents tricuspidées, caractéristiques des
Nesophontidae, le distinguaient nettement des Soricidae
modernes. Il avait probablement une fourrure dense, une
queue modérément longue et des membres courts adaptés à une
vie fouisseuse dans la litière forestière. Sa morphologie
générale suggère un animal discret, terrestre, et spécialisé
dans la recherche d’invertébrés au sol.
L’écologie du Nesophonte de Porto Rico est déduite de ses
caractéristiques dentaires et de ses proches parents. Il
était très probablement
insectivore,
se nourrissant d’insectes, d’arthropodes, de larves, de vers
et peut‑être de petits invertébrés du sol. Il vivait dans la
litière forestière,
sous les rochers, dans les crevasses et les zones humides
des forêts tropicales. Il devait être
nocturne
ou
crépusculaire,
actif au sol, et jouait un rôle écologique important dans le
contrôle des populations d’insectes. Les fossiles montrent
qu’il cohabitait autrefois avec des oiseaux endémiques, des
reptiles et d’autres petits mammifères, avant l’arrivée des
espèces introduites par l’homme.
La disparition de
Nesophontes portoricensis
est étroitement liée à l’arrivée des
Européens
dans les Caraïbes et à l’introduction de
prédateurs exotiques,
en particulier la
ratte noire
(Rattus
rattus)
et la
ratte brune
(Rattus
norvegicus).
Ces rongeurs, arrivés dès le XVIᵉ siècle, ont provoqué une
prédation directe,
une
compétition alimentaire,
et probablement une
transmission de maladies
auxquelles les Nesophontes n’étaient pas adaptés. La petite
taille de l’île et la fragmentation des habitats ont aggravé
la situation. La
déforestation,
les
incendies,
la
modification des milieux forestiers
et l’expansion des plantations ont réduit les zones de
refuge. Les études paléontologiques montrent que l’espèce a
survécu jusqu’à une période très récente, probablement
jusqu’au
XIXᵉ siècle,
voire au début du
XXᵉ siècle,
avant de disparaître complètement. L’UICN
classe aujourd’hui l’espèce comme
éteinte (EX).
Sa disparition s’inscrit dans un effondrement plus large des
petits mammifères endémiques des Antilles, dont toutes les
espèces du genre
Nesophontes
ont disparu en quelques siècles seulement, principalement à
cause des espèces introduites. |
|
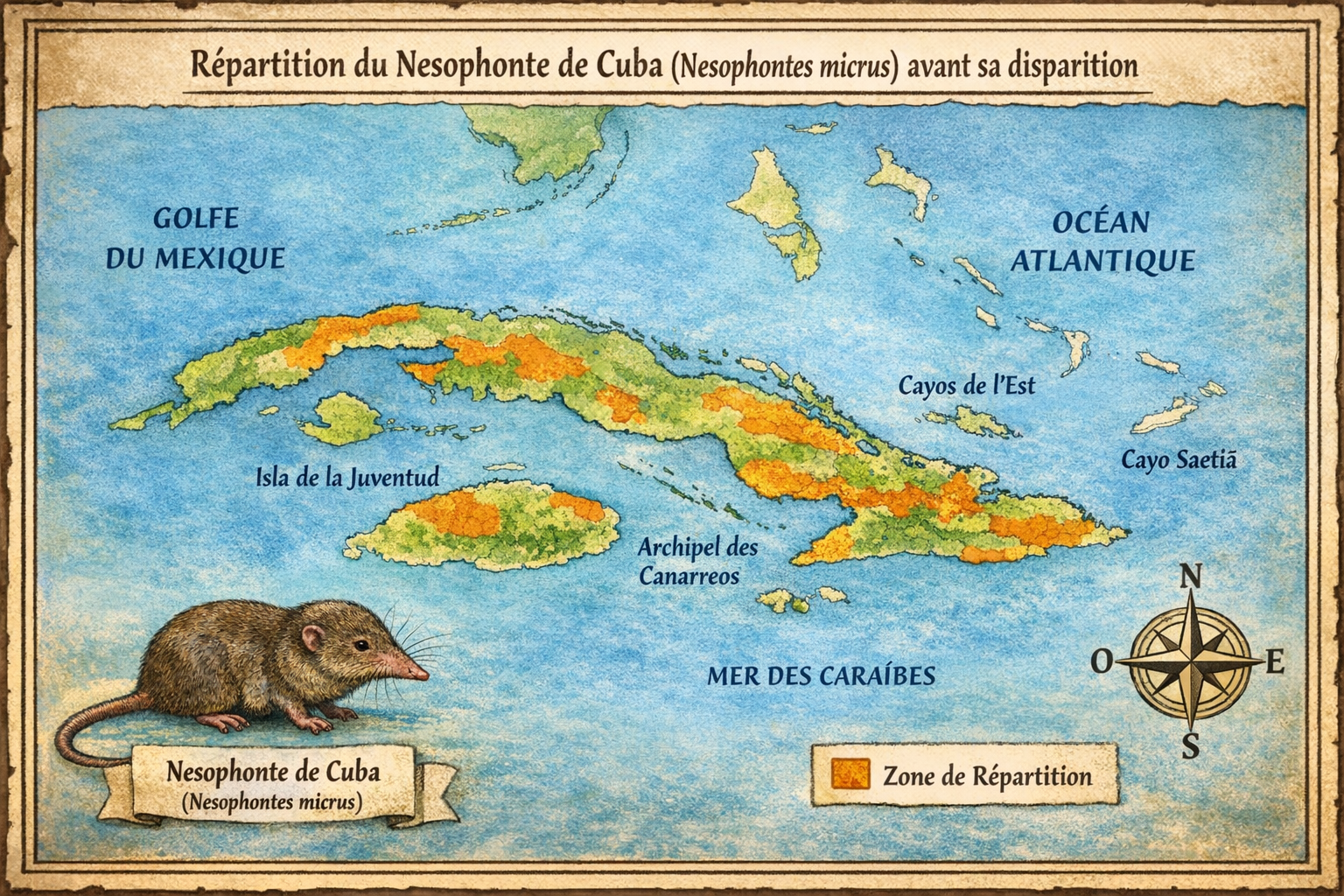
Nesophonte de Cuba (Nesophontes micrus)

 |
EX
|
Le
Nesophonte de Cuba
(Nesophontes
micrus)
appartenait à la famille éteinte des
Nesophontidae,
un groupe de petits insectivores endémiques des
Antilles.
Cette espèce était strictement
endémique de Cuba,
où elle occupait autrefois une grande partie de l’île
principale ainsi que certaines zones karstiques et
forestières. Les fossiles montrent qu’elle était présente
depuis le
Pléistocène tardif,
et qu’elle cohabitait avec d’autres espèces du genre
Nesophontes
aujourd’hui disparues. Sa répartition historique couvrait
les
forêts humides,
les
forêts sèches,
les
zones rocheuses,
les
grottes
et les
milieux de litière forestière,
ce qui en faisait l’un des Nesophontes les plus largement
distribués dans les Grandes Antilles.
Nesophontes micrus
était un petit mammifère insectivore, de taille comparable à
une
musaraigne,
avec un corps d’environ
8 à 10 cm
et un poids estimé entre
15 et 30 g,
ce qui en faisait l’un des plus petits membres du genre. Il
possédait un
museau long et étroit,
des
dents fines et tricuspidées
adaptées à la capture d’insectes, et un crâne
proportionnellement robuste. Ses membres courts et sa queue
modérée suggèrent un animal terrestre, fouisseur dans la
litière forestière. Sa morphologie générale rappelle celle
des Soricidae, mais les Nesophontidae formaient une lignée
totalement distincte, isolée dans les Antilles depuis
plusieurs millions d’années.
L’écologie de
Nesophontes micrus
est déduite de ses caractéristiques dentaires et de ses
proches parents. Il était très probablement
insectivore,
se nourrissant d’insectes, d’arthropodes, de larves, de vers
et de petits invertébrés du sol. Il vivait dans la
litière forestière,
sous les rochers, dans les crevasses et les zones humides
des forêts tropicales cubaines. Il devait être
nocturne
ou
crépusculaire,
actif au sol, et jouait un rôle écologique important dans le
contrôle des populations d’insectes. Les fossiles montrent
qu’il cohabitait autrefois avec des oiseaux endémiques, des
reptiles et d’autres petits mammifères avant l’arrivée des
espèces introduites par l’homme.
La disparition de
Nesophontes micrus
est étroitement liée à l’arrivée des
Européens
dans les Caraïbes et à l’introduction de
prédateurs exotiques,
en particulier la
ratte noire
(Rattus
rattus)
et la
ratte brune
(Rattus
norvegicus).
Ces rongeurs, arrivés dès le XVIᵉ siècle, ont provoqué une
prédation directe,
une
compétition alimentaire,
et probablement une
transmission de maladies
auxquelles les Nesophontes n’étaient pas adaptés. La
déforestation,
les
incendies,
la
modification des habitats
et l’expansion des plantations ont réduit les zones de
refuge. Les études paléontologiques montrent que l’espèce a
survécu jusqu’à une période très récente : des restes datés
du
XIXᵉ siècle,
voire du
début du XXᵉ siècle,
suggèrent qu’elle a disparu seulement quelques décennies
avant sa description scientifique. L’UICN
la classe aujourd’hui comme
éteinte (EX).
Sa disparition s’inscrit dans un effondrement plus large des
petits mammifères endémiques des Antilles, dont toutes les
espèces du genre
Nesophontes
ont disparu en quelques siècles seulement, principalement à
cause des espèces introduites. |
|